Мезофиты.
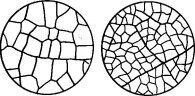
Рис. 106. Нижний эпидермис и густота жилкования в листьях ясеня. А — в лесу; Б — на полезащитной полосе
Основные морфолого-анато- мические и физиологические черты мезофитов — средние между чертами гигрофитов и ксерофитов, так называемые мезофиль- ные. Мезофиты имеют умеренно развитые корневые системы как экстенсивного, так и интенсивного типа, со всеми переходами между ними. Для листа характерна дифференцировка тканей на более или менее плотную палисадную паренхиму (за исключением листьев ярко выраженного теневого характера) и рыхлую губчатую паренхиму с системой межклетников. Сеть жилок сравнительно негустая. Покровные ткани могут иметь отдельные ксероморфные черты (чаще всего редкое опушение), но не столь ярко выраженные, как у ксерофитов.
Физиологические показатели водного режима мезофитов (см. с. 110—119, табл. 10, 11) подтверждают их промежуточную пози-
Таблица 12. Различие анатомо-физиологических показателей листьев иа разных сторонах кроны дерева, растущего иа опушке леса.
1—сторона, обращенная к лесу, 11 —сторона, обращенная к поляне
| Древесная порода | Площадь листа, см® | Число устьиц на 1 мм2 | Содержание во- ДЫ, % | Содержание хлорофилла, мг/г |
Средняя интенсивность фотосинтеза ,
•мг С02/дм2-ч |
Толщина листа, мкм | |
| Дуб — I | Quercus robur | 42 | 45 | 61 | 3,0 | 1,9 | 97 |
| 11 | 18 | 125 | 54 | 2,4 | 2,3 | 181 | |
| Липа — I | Tilia cordata | 38 | 38 | 71 | 3,6 | 1,6 | 93 |
| II | 24 | 45 | 62 | 2,0 | 1,1 | 106 |
цию; для них характерны умеренные величины осмотического давления, содержания воды в листьях, предельного водного дефицита.
Что касается транспирации, то ее величина в большой степени зависит от условий освещенности и других элементов микроклимата; у луговых растений она больше, под пологом лесов — гораздо - меньше.Один и тот же мезофильный вид, попадая в разные по водоснабжению условия, обнаруживает известную пластичность, приобретая в сухих условиях более ксероморфные, а во влажных — более гигроморфные черты. Это хорошо иллюстрируют рис. 106 и 107, показывающие, что с усилением сухости и освещенности у мезофитов наблюдается не только определенная ксерофилизация листа, но изменяется и интенсивность физиологических процессов (водного режима, дыхания и др.).
Пластичность листьев проявляется не только в разных местообитаниях, но даже у одной и той же особи. Например, у деревьев
| т
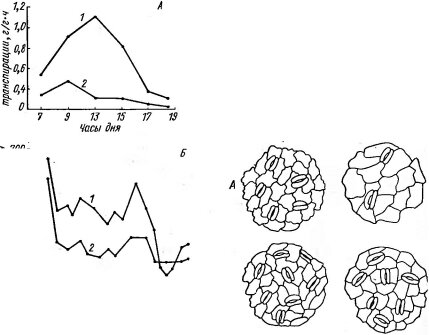
Рис. 108. Нижний эпидермис листьев на разных сторонах кроны дерева, растущего на опушке леса. А — сторона дерева, обращенная к лесу; Б — сторона, обращенная к поляне; 1 — дуб — Quercus robur, 2 — липа — Tilia cordata
? 600 I
^500
g 300
Со
1 200 g
? wo §
IV V VI VII VIII IX Месяцы
на опушке леса листья на стороне, которая обращена в сторону леса, имеют более мезофильный и теневой характер по сравнению с несколько ксе- роморфными листьями внешней стороны дерева, (рис. 108, табл. 12). Листья разных высотных ярусов одних и тех же
Рис. 107. Изменение интенсивности физиологических процессов у травянистых растений- мезофитов в местообитаниях с различным микроклиматом. А — дневной ход транспирации у сныти — Aegopodium podagraria; Б — сезонный ход дыхания листьев у ежи — Dactylis glomerata:
I — на открытой лесной поляне, 2 — под пологом
дубового леса
растений (как травянистых, так и особенно древесных) находятся в неодинаковых условиях водоснабжения, так как поступление воды в верхние части связано с преодолением большего сопротивления (силы тяжести, трения).
К тому же у деревьев верхние листья обычно находятся в условиях иного микроклимата (большее осве-1 г з
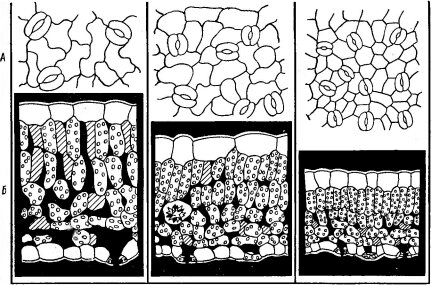
Рис. 109. Различие анатомического строения листьев разных ярусов у таволги Filipendula ulmaria (по Yapp R. Н., 1912). А — клетки нижнего эпидермиса с устьицами; Б — поперечный срез листа.
Листья. / — нижний, 2 — средний, 3 — верхний
щение, нагревание и иссушение, ветер). Поэтому, как правило, верхние листья крупных растений имеют более ксероморфный характер, чем средние и тем более нижние (закон В. Р. Заленского, рис. 109).
Мезофиты — группа весьма разнообразная не только по видовому составу, но и по различным экологическим «оттенкам», обусловленным разным сочетанием факторов в природных местообитаниях. Они связаны переходами с другими экологическими типами растений по отношению к воде, так что четкую границу между ними провести очень трудно. Так, среди луговых мезофитов выделяются виды с повышенным влаголюбием, предпочитающие постоянно сырые или временно заливаемые участки (лисохвост луговой— Alopecurus pratensis; канареечник тростниковидный — Dig- raphis arundinacea; бекмания обыкновенная — Beckmannia erucifor- mis и др.). Их объединяют в переходную группу гигромезофитов наряду с некоторыми влаголюбивыми лесными травами, предпочитающими наиболее сырые леса, лесные овраги и т. д. (селезеночник — Chrysosplenium alternifolium, недотрога — Impatiens noli tangere, некоторые лесные мхи и папоротники). С другой стороны, в местообитаниях с периодическим или постоянным (но небольшим) недостатком влаги.много мезофитов с теми или иными ксеро- морфными признаками или с повышенной физиологической устойчивостью к засухе. Эта группа, переходная между мезофитами и ксерофитами, — ксеромезофиты. Примером могут служить многие виды северных («луговых») степей, сухих сосновых боров, песчаных местообитаний: клецер-белоголовка — Trifolium monta- пит, подмаренник желтый — Galium verum, и др.
Из культурных растений к ксеромезофитам можно отнести люцерну, засухоустойчивые сорта пшениц. Близки к этой группе доминирующие травянистые виды лесостепных дубрав (сныть, копытень, ясменник и др.). Несмотря на вполне мезофильный внешний облик, они способны переносить длительные засухи и состояние завядания.Особое место среди мезофитов занимают степные и пустынные весенние эфемеры и эфемероиды. К этой группе принадлежат растения, ранней весной покрывающие степи и пустыни разноцветным цветущим ковром (многолетники — тюльпаны, гусиные луки, птицемлечники, гадючий лук; однолетники — крупки, маки, вероники и др.). Это виды с чрезвычайно краткой вегетацией (иногда не более 4—6 недель) и длительным периодом покоя, который однолетние эфемеры переживают в виде семян, а многолетние эфемероиды — в виде покоящихся луковиц, клубней, корневищ. На первый взгляд эти растения трудно отнести к мезофитам, поскольку они в основном распространены в областях жаркого и сухого климата. Однако благодаря очень ранним срокам вегетации эфемерам и эфемероидам удается избежать засухи. Сезонное развитие у степных видов начинается сразу после снеготаяния, а у пустынных эфемеров (в случае мягкой зимы)—еще раньше[V], и заканчивается в конце весны или самом начале лета, еще до наступления сильной жары и засухи; затем следует длительный летний покой. Благодаря такому сдвигу сезонного цикла развития по отношению к обычному климатическому ритму умеренных широт степные и пустынные эфемеры и эфемероиды занимают особую сезонную экологическую нишу, хорошо обеспеченную влагой, и принадлежат к мезофитам. Многие имеют крупные сочные листья (тюльпаны, виды из родов ревень — Rheum, смолоносница — Ferula) , совершенно немыслимые у пустынных растений летом, а также крупные околоцветники (также испаряющие много влаги). Транспирация (см. выше) у эфемеров и эфемероидов, несмотря на низкие температуры ранней весны, в несколько раз выше, чем у ксерофитов, вегетирующих летом на тех же территориях. Другие показатели водного режима (см.
с. 111—119, табл. 10) отражают вполне достаточное водоснабжение и отсутствие необходимости защиты от иссушения.Кроме весенних, существуют и осенние эфемероиды (виды родов Crocus, Scilla и др.), произрастающие в районах с климатическим ритмом средиземноморского типа (летней засухой и осенне-зимними осадками). Это тоже растения вполне мезофильной организации.
К мезофитам принадлежат и ранневесенние эфемероиды лиственных лесов (см. с. 52). У этой группы сдвиг вегетации на раннюю весну обусловлен не столько условиями увлажнения, сколько сезонной динамикой освещенности под пологом леса. Вегетируя в условиях изобилия почвенной влаги после снеготаяния, лесные эфемероиды имеют не только типично мезофильные, но отчасти и гигрофильные черты.
По многим особенностям структуры и физиологии близки к ксерофитам растения, которые по тем или иным причинам испытывают недостаток влаги, сопряженный с действием низких температур. Иногда такие виды в качестве особого подразделения включают в группу ксерофитов, иногда выделяют в самостоятельные экологические типы — психрофиты и криофиты.
Психрофиты — растения влажных и холодных почв в холодных местообитаниях высокогорий и северных широт. Несмотря на достаточное увлажнение почвы, они часто испытывают недостаток влаги (или из-за физиологической сухости, вызванной низкими температурами, или в связи с преобладанием в почве недоступной влаги, как, например, на торфянистых почвах). В отдельные периоды возможна и физическая сухость. Среди психрофитов есть травянистые растения (например, злаки северных лугов: белоус — Nar- dus stricta, душистый колосок — Anthoxanthum odoratum, луговик извилистый — Deschampsia flexuosa; высокогорные кавказские злаки: овсяница пестрая — Festuca varia и др., осоки, виды рода КоЪ- resia), высокогорные, болотные и тундровые кустарники и кустарнички, как вечнозеленые (дриада — Dryas octopetala, брусника— Vaccinium vitis idaea, водяника— Empetrum nigrum, вереск— Cal- luna vulgaris, кедровый и можжевеловый стланики), так и с опадающей листвой (карликовые ивы — Salix polaris, S.
herbacea, голубика— Vaccinium uliginosum и др.). К психрофитам относятся и хвойные древесные породы умеренных и северных широт. Из низших растений психрофитами являются водоросли, обитающие на снегу и льду, наскальные и напочвенные тундровые лишайники.Все психрофиты имеют ярко выраженную ксероморфную структуру листа. Так, психрофильные злаки узколистны, имеют хорошо развитую проводящую и механическую ткани; некоторые из них способны к свертыванию листовых пластинок в трубку, напоминая степные ксерофиты. Вечнозеленые кустарнички имеют плотные кожистые листья, иногда с весьма мощной кутикулой, плотной палисадной паренхимой. Нижняя сторона, несущая устьица, часто опушена (багульник — Ledum palustre) или покрыта восковым налетом (подбел — Andromeda polifolia). Вместе с тем в структуре листа ясно заметны и некоторые гигроморфные черты; главным образом, крупные размеры клеток и развитие больших межклетников в губчатой ткани. Этн особенности отличают листья психрофитов от вечнозеленых ксерофитов (лавров, олеандров и т.д.). Своеобразно строение листа у водяники, вереска и родственных ему растений из
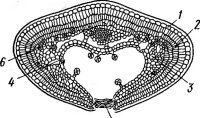
5
Рис. 110- Строение листа водяники Empetrum nigrum:
семейства Ericaceae (так называемая эрикоидная структура): листья мелкие, игловидные, но с рыхлой паренхимой. Как видно на рис. 110, благодаря тому что края листа сильно загнуты вниз, нижняя поверхность, несущая устьица, открывается в полость, вход в которую закрыт волосками.
I — верхний эпидермис с толстой кутикулой, 2 — палисадная ткань, 3 — губчатая ткань, 4 — нижний эпидермис с устьицами
Особо следует отметить ксеро- фильные черты в строении игл хвойных древесных пород, известные из курса анатомии растений (толстостенный эпидермис с под- и стилающей его гиподермой, погру
женные устьица, закрываемые зимой смоляными пробками, ит. д.). Здесь ксероморфоз, возможно, связан не только с условиями обитания, но и с трудностями подачи воды к высоко расположенным кронам через систему трахеид (а не сплошных сосудов, как у покрытосеменных). Ксероморфные черты хвои особенно необходимы зимой, так как предотвращают лишнее испарение в условиях, когда корни почти не всасывают влаги.
Водный режим психрофитов в целом сформировался под влиянием затрудненного поступления воды, а также других неблагоприятных условий северных и высокогорных местообитаний (сильных ветров, низких температур и т. д.). Расход воды на транспирацию у психрофитов относительно невелик (что особенно хорошо видно при сопоставлении транспирации хвойных и лиственных древесных пород, см. с. 114) и способность выносить большие потери воды гораздо меньше, чем у настоящих ксерофитов. По основным физиологическим показателям водного режима психрофиты представляют весьма неоднородную группу: так, растения сравнительно влажных высокогорных областей (Кавказ, Альпы) ближе к мезофитам, чем психрофиты сухих высокогорий или тундры.
Криофиты в экологическом отношении очень близки к псих- рофитам и связаны с ними переходными формами. Это растения сухих и холодных местообитаний — сухих участков тундр, скал, осыпей и т. д. Обычно они рассматриваются и характеризуются вместе с психрофитами, поскольку у них много сходных морфологических и физиологических черт. Но среди криофитов есть и весьма своеобразные формы — это растения-подушки высокогорных холодных пустынь (см. с. 86). Как показали исследования на Памире, для водного режима подушек характерны замедленные темпы водообмена, низкая оводненность тканей и весьма экономное . расходование воды.
Еще по теме Мезофиты.:
- Экологические группы растений по отношению к влажности.
- О ПОНЯТИИ «ЖИЗНЕННАЯ ФОРМА»
- География растений
- Индикаторы грунтовых вод
- РАСТЕНИЕ И РАСТИТЕЛЬНЫЙ ПОКРОВ КАК ОПТИЧЕСКАЯ СИСТЕМА
- Портал "ПЛАНЕТА ЖИВОТНЫХ". Кто ты, собака?, 2010
- Любопытное доказательство того, что собаки очень давно одомашнены, приводит советский ученый-языковед академик Н. Я. Марр...
- Антропологи изучают кости и скелеты людей очень далекого прошлого, изучают их близких и отдаленных родственников — ископаемых и современных обезьян,— чтоб восстановить путь, который прошел человек в своем развитии.
- Находки, проливающие свет на происхождение собак, имеют возраст 8—10 тысяч лет...
- Значит, собака пришла сюда вместе с человеком?..
- Находка Савенкова произвела сенсацию...