Характеристика подтипа
Более высоко организованные хордовые животные. Отличаются активным питанием: пища разыскивается, нередко преследуется, а после поимки часто измельчается, что облегчает переваривание.
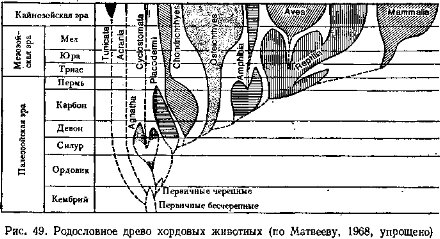
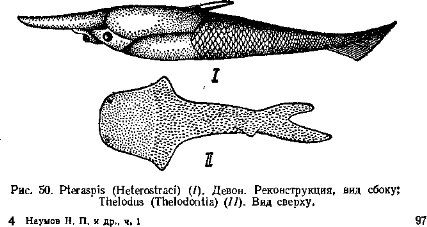
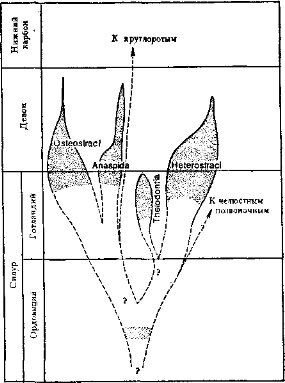
Переход к активному питанию сопровождался усилением подвижности и перестройкой двигательной системы, хотя ее принципиальная схема сохраняется. Хорда замещается позвоночником, развивается череп (защита головного мозга), вооруженный челюстями (органом захвата и измельчения пищи); возникают парные конечности и их пояса. Уровень метаболизма у позвоночных существенно повышается за счет интенсификации питания, пищеварения, дыхания, кровообращения и выделения. Существенно усложняется строение и функции центральной нервной системы, органов чувств и гуморальной (гормональной) регуляции. На этой основе усложняется поведение и популяционная (внутривидовая) организация. Растущую роль в жизни позвоночных начинают играть группировки особей (семьи, стаи, колониальные поселения и др.), упорядочивающие использование среды, увеличивающие возможности питания, эффективность размножения и снижающие смертность. Все это объясняет явную тенденцию к падению индивидуальной плодовитости в ряду позвоночных. Рост уровня жизнедеятельности, совершенствование размножения и заботы о потомстве сопровождалось расселением позвоночных по всей поверхности Земли: в морях и океанах, пресных водоемах и на суше — от тропиков до высоких широт Арктики и Антарктики.Подтип позвоночных разделяют на два раздела (см. с. 12): бесчелюстные — Agnatha, который включает два вымерших класса, и современный класс круглоротые, и челюстноротые — Gnathostomata, которые объединяют два надкласса: рыбы —Pisces (включает два ископаемых и два современных класса) и четвероногие — Tetrapoda с четырьмя классами — земноводные, пресмыкающиеся, птицы и млекопитающие. Челюстноротые обычно подразделяются на две группы, не имеющие таксономического значения: первичноводные — Anamnia (рыбы и земноводные; последние вышли на сушу, но сохранили тесные связи с водой) и первичноназемные — Amniota (пресмыкающиеся, птицы, млекопитающие; некоторые из них вторично перешли к водному образу жизни).
Наконец, различия в устойчивости метаболизма и степени стабильности внутреннего климата отличают пойкилотермных (холоднокровных или экзотермных) позвоночных — круглоротых, рыб, земноводных и пресмыкающихся — от гомойотермных (теплокровных или эндотермных) позвоночных — птиц и млекопитающих. В современной фауне к подтипу позвоночные относятся около 42 тыс. видов (Майр, 1971).Основные черты организации позвоночных
Форма тела. Водные позвоночные обычно имеют «рыбообразное» веретеновидное удлиненное тело, подразделяемое на голову, туловище и хвост. Развиваются парные (отсутствуют у круглоротых) и непарные плавники. Форма тела наземных позвоночных более разнообразна. У них появляется шейный отдел, увеличивающий подвижность головы; во многих группах уменьшается хвостовой отдел. Непарные плавники редуцируются, а парные превращаются в рычажные конечности наземного типа, у китообразных и ихтиозавров (вымерших рептилий) преобразующиеся в плавники.
Кожные покровы. Кожа позвоночных животных двухслойна: наружный слой — многорядный эпидермис, внутренний кориум, или собственно кожа. Эпидермис развивается из наружного зародышевого листка — эктодермы — и состоит из многих рядов клеток, из которых нижний представлен энергично размножающимися клетками (мальпигиев слой), восстанавливающими нарушаемые поверхностные слои. В эпидермальном слое имеются различного назначения железистые клетки, у наземных позвоночных часто образующие многоклеточные железы, которые погружаются в слой кориума. В эпидермальном слое возникают защитные образования: эмалевые части плакоидных чешуй, роговые чешуи пресмыкающихся, когти и перья птиц, когти и волосы млекопитающих. Скопление пигмента в клетках эпидермиса обусловливает окраску кожи (хроматофорные клетки).
Кориум развивается из наружного листка миотомов зародыша, т. е. из мезодермы. Обычно он толще слоя эпидермиса и образуется путем разрастания волокнистой соединительной ткани, пронизанной кровеносными сосудами и нервными окончаниями.
В этом слое возникают покровные скелетные образования: костные чешуи и покровныё кости. Скопления пигмента концентрируются в отдельных пигментных клетках и обусловливают вместе с пигментами эпителиального слоя специфичную для каждого вида окраску, подчас очень яркую и сложную (имеет важное сигнальное значение).Помимо функции механической защиты подлежащих тканей кожа участвует в обмене веществ (водном и солевом обмене, дыхании и др.), в терморегуляции и иных физиологических процессах. Ее особо важная роль — защита организма от проникновения болезнетворных агентой с помощью механизмов фагоцитарной и биохимической природы.
Скелет. Существенно дифференцируется скелет. Он представлен осевым скелетом, черепом, скелетом конечностей’ и их поясов. У бесчелюстных и хрящевых рыб он построен разными типами хряща. Иногда хрящ импрегнируется кальциевыми солями и принимает внешне вид кости. У остальных позвоночных внутренний скелет в основном костный с некоторым участием хряща.
В эволюционном ряду позвоночных кость вначале появилась в покровных образованиях (костные шипы и щитки бесчелюстных, панцирные пластинки и костная чешуя у рыб). Она служила защитой от механических повреждений и врагов. У пресноводных обитателей костный покров уменьшал обводнение организма, грозившее наруше-
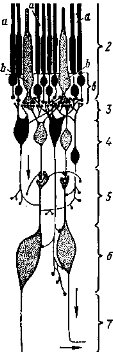

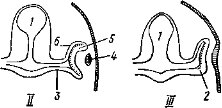 Рис. 30. Гистологическое строение хряща. А — хрящ круглоротых (межклеточное вещество слабо развито); Б — гиалиновый хрящ челюстноротых (хорошо развито межклеточное вещество) (по Румянцеву, 1958)
Рис. 30. Гистологическое строение хряща. А — хрящ круглоротых (межклеточное вещество слабо развито); Б — гиалиновый хрящ челюстноротых (хорошо развито межклеточное вещество) (по Румянцеву, 1958)
нием нормального осмотического давления внутренней среды. Предполагают, что в эволюции скелета исходной тканью был «слизистый хрящ», малопрочный и слабоупругий, подобно встречающемуся в скелетах личинок круглоротых. Позднее в скелетных образованиях появились упругие, эластичные и высокопрочные гиалиновые и волокнистые хрящи (рис.
30) и, наконец, возникла настоящая кость разного строения и прочности: от грубоволокнистой у рыб до тонковолокнистой пластинчатой кости со сложной остеонной организацией у рептилий, птиц и млекопитающих (А. В. Румянцев, 1958) х.Таким образом, в ходе эволюции внутреннего скелета позвоночных соответственно среде обитания и образу жизни усложнялось и совершенствовалось не только его анатомическое строение, но и гистологическая структура материала, из которого построен скелет. На этом примере можно видеть, как эволюционные преобразования на уровне организма (анатомия) сочетаются с изменениями на клеточно-молекулярном уровне. Важно заметить, что оба ряда изменений взаимосвязаны, но и относительно независимы, т. е. не строго синхронны.
В онтогенезе позвоночных скелетные элементы возникают в волокнистой соединительной ткани (вероятно, представляющей преобразованную слизистою опорную ткань бесчелюстных) в виде хрящевых образований, которые затем могут замещаться костью (замещающие, хрящевые или хондральные кости). Другие кости образуются в соединительнотканном слое кожи сразу, не проходя хрящевой стадии, — это покровные или кожные кости. Нередко они погружаются под кожу и срастаются с хрящевыми костями. По внешнему виду или положению определить тип и происхождение кости невозможно, оно устанавливается лишь при изучении ее развития. />В эволюционном ряду позвоночных животных хорда замещается позвоночным столбом, состоящим из хрящевых или костных позвонков. Они образуются в соединительнотканной оболочке хорды. У кругло-
J Подробнее о микроструктуре костей см. в описаниях скелетов разных классов.
61
ротых хорда полностью сохраняется, но в ее соединительнотканной оболочке развиваются палочковидные хрящи, которые рассматриваются как зачатки верхних дуг позвонков. У большинства рыб хорда, в большей или меньшей степени сжатая и деформированная образовавшимися телами хрящевых или костных позвонков, все же сохраняется, но опорой не служит. В остальных классах позвоночных животных хорда развивается лишь у зародышей, а затем вытесняется позвонками.
Замещение хорды сегментарным позвоночным столбом увеличивает прочность осевого скелета при сохранении его гибкости (подвижности). Сложный рельеф позвонков обеспечивает возможности прикрепления мощной мускулатуры. Верхние дуги позвонков образуют канал, в котором лежит спинной мозг.Зачатки парных и непарных конечностей в виде примитивных стабилизаторов — метаплевральных и непарной плавниковой складок — наметились еще и у бесчерепных. У водных позвоночных на их основе развились парные (грудные и брюшные) и непарные (хвостовой, спинной, анальный) плавники с их внутренним и наружным скелетом. У наземных позвоночных парные плавники рыбообразных предков превратились в членистые конечности, в зависимости от способов передвижения приобретающие форму лап, ног, ласт или крыльев. Скелет парных конечностей крепится с помощью переднего или грудного и заднего или тазового поясов конечностей. Они по-разному соединяются с осевым скелетом, соответственно характеру движения.
Мускулатура. Позвоночный столб, скелет конечностей и их поясов служат опорой и местом прикрепления двигательной мускулатуры, мощность которой у позвоночных по сравнению с низшими хордовыми резко возрастает. У водных позвоночных двигательная мускулатура имеет метамерное строение, что обусловлено участием большей части тела в двигательном акте. Метамерность нарушается лишь в области парных плавников, где путем слияния и дифференцировки брюшных участков миомеров формируются глубокие и поверхностные мышцы плавников. Не имеет метамерного строения и мускулатура висцерального черепа, приводящая в движение челюсти и жаберный аппарат. С переходом к наземному образу жизни, в связи с опорой преимущественно на парные конечности метамерность двигательной мускулатуры существенно нарушается: участки отдельных миомеров сливаются друг с другом, образуя лентовидные и иной формы мышцы (мускулы), лежащие в нескольких сегментах тела; возникают мощные и сложные мускульные комплексы сгибателей и разгибателей. При этом возрастает масса мускулатуры конечностей, которая частично размещается и на туловище.
Одновременно собственно туловищная мускулатура уменьшается. У высших позвоночных метамерность проявляется лишь в расположении мелких мускулов позвоночного столба, части мышц брюшного пресса и межреберных мышц. Все эти изменения в скелете и мускулатуре обеспечивают значительное возрастание подвижности и маневренности позвоночных животных.Центральная нервная система. Одновременно с совершенствованием органов движения развивалась центральная нервная система с ее периферией, управляющая сложным комплексом движений орга
низма и регулирующая его физиологические функции. Эти функции выполняются на основе анализа сигналов, воспринимаемых органами чувств. На той же базе осуществляется ориентация животного в пространстве и формирование его поведения в меняющейся среде.
 Закладывающаяся на спинной стороне зародыша позвоночных нервная пластинка (рис. 31) погружается под кожу и сворачивается в трубку, передний конец которой открывается отверстием — невро- пором — наружу, а задний — ней- рокишечным каналом соединяется с полостью первичной кишки. Оба отверстия вскоре закрываются.
Закладывающаяся на спинной стороне зародыша позвоночных нервная пластинка (рис. 31) погружается под кожу и сворачивается в трубку, передний конец которой открывается отверстием — невро- пором — наружу, а задний — ней- рокишечным каналом соединяется с полостью первичной кишки. Оба отверстия вскоре закрываются.
При замыкании трубки нервные складки образуют в ее верхней части парные боковые выросты — ганглионарные пластинки, из которых в дальнейшем образуются спинномозговые узлы, часть ганглиев головных нервов и ганглии симпатической (вегетативной) нервной системы.
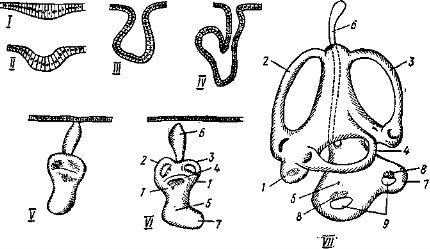
Далее нервная трубка дифференцируется на головной и спинной мозг. В ее передней части возникает небольшое вздутие, из которого последовательно образуются три мозговых пузыря: передний, средний и задний (рис. 32). Дальнейшая дифференцировка приводит к образованию пяти отделов головного мозга. Передняя часть переднего пузыря, разрастаясь, дает собственно передний мозг (telencephalon), который у большинства позвоночных, разделяясь продольной складкой, образует обособленные полушария мозга; заключенные в них полости называются боковыми желудочками. Задняя часть переднего пузыря превращается в промежуточный мозг (diencephalon), образующий парные боковые выпячивания — глазные пузыри, из которых в дальнейшем формируются сетчатка и пигментная оболочка глаза (см. ниже); полость промежуточного мозга получает название третьего желудочка. Средний пузырь превращается в средний мозг (mesencephalon), а его полость называется сильвиевым водопроводом. Передняя часть заднего пузыря образует выступающий вверх задний мозг, или мозжечок (cerebellum). Остальная часть заднего пузыря разрастается в продолговатый мозг (myelencephalon), без резкой границы переходящий в спинной мозг. Полость продолговатого мозга — четвертый желудочек, или ромбовидная ямка, — продолжается в полость спинного мозга — невроцель. Эти пять отделов головного мозга характерны для всех позвоночных животных, но степень их развития, детали строения и функции существенно отличаются в разных классах.
Нервные клетки позвоночных в общем сходны с нейронами других животных, но имеют свои особенности; их тела и короткие отростки — дендриты — составляют серое вещество мозга, а длинные отростки нервных клеток — аксоны, или невриты, окруженные неврилеммой
18
Рис. 32. Схема развития головного мозга позвоночных (по Паркеру, с изменен.): I — первая стадия (сбоку); II — вторая стадия (сбоку); III — развитый мозг с неразделенным передним мозгом; IV — то же, сагиттальный разрез; V — головной мозг с парными полушариями; VI — то же, сагиттальный разрез; VII — то же, сверху (правая сторона переднего и среднего мозга вскрыты):
/ — передний мозг, 2 — средний мозг, 3 — задний мозг, 4 — спиниой мозг, 5 — промежуточный мозг, 6 — мозжечок, 7 — продолговатый мозг, 8 — обонятельная доля, 9 — гипофиз, 10 — париетальный орган, II — эпифиз, 12 — желудочек переднего мозга, 13 — зрительный бугор, 14 — третий желудочек, 15 — желудочек среднего мозга, 16 — желудочек мрзжечка, 17 — четвертый желудочек, 18 — полушарие переднего мозга, 19 — канал спинного мозга, 20 — желудочек обонятельной доли, 21 — правый боковой желудочек, 22 — полосатое тело, 23 — зрительная доля (двухолмие)
(шванновскими клетками) и миэлиновой оболочкой, образуют белое вещество (рис. 33). Оболочка изолирует нервные волокна, обеспечивая независимость проведения импульса (подобно изоляции электрических проводов). В организме позвоночного животного, где одновременно получается, передается и обрабатывается огромное количество разнообразной информации, подобное усложнение нервных путей необходимо.
Основная масса серого вещества переднего мозга расположена в виде полосатых тел на дне и в его парных выпячиваниях — обонятельных долях. От каждой обонятельной доли отходит обонятельный тракт, образующий у обонятельной капсулы расширение — луковицу. Короткий обонятельный нерв (nervus olfactorius) (I пара головных нервов) состоит из чувствующих волокон, связывающих клетки эпителия обонятельной капсулы с луковицей. Тонкий слой серого вещества выстилает стенки полостей желудочков. Верх переднего мозга — мантия — у большинства низших позвоночных образован только белым веществом; лишь у двоякодышащих рыб и земноводных в ее поверхностном слое появляется небольшое число нервных клеток. У пресмыкающихся в мантии имеется серое вещество (скопление нервных клеток), представляющее зачаток коры больших полушарий. У птиц
Ill
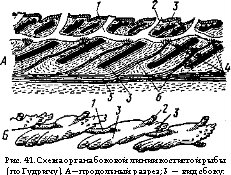
Рис. 33. Три типа нейронов (Л) н этапы формирования оболочек аксона (Б) (по Гриффину и Новику, 1973): I—двигательный нейрон (мотонейрон); II — чувствительный (рецепторный) нейрон; III — нейрон центральной
нервной системы:
/ — деидриты, 2 — тело клетки, 3 — ядро, 4 — цитоплазма, 5 — аксои, 6 — шваииовская клетка, 7 — окончания аксона, 8 — аксои, 9 — шпаниовская клетка, 10 — оболочка аксона и шваиновской клетки, 11 — щель, 12 — собственная оболочка аксона, 13 — цитоплазма шваиновской клетки
объем переднего мозга резко возрастает, но сохраняется тот же тип строения, что и у пресмыкающихся. У млекопитающих величина переднего мозга увеличивается не столько за счет разрастания полосатых тел, сколько путем развития серого вещества на поверхности мантии. Этот слой нервных клеток, функционирующий как высший ассоциативный центр, называют корой больших полушарий. Его подразделяют на первичную кору, или гиппокамп (archipallium, seu hipocampus) — разрастание образовавшегося у пресмыкающихся зачатка коры, и на вторичную кору (neopallium). На ней образуются  Наумов Н. П. н Др., ч. 1
Наумов Н. П. н Др., ч. 1
борозды (извилины), за счет которых возрастает наружная поверхность коры.
 Утолщенные стенки промежуточного мозга называют зрительными буграми (thalami optici) (см. рис. 32,13)\ выросты их стенок образуют зрительные нервы (п. optici) (II пара головных нервов), имеющие хиазму (перекрест): часть волокон нерва правой стороны уходит в левый нерв, а часть волокон слева переходит в правый нерв. Зрительные нервы иннервируют только сетчатку глаза. Позади хиазмы на дне промежуточного мозга образуется полый вырост — воронка (infundibulum), к передней стенке которого примыкает железа внутренней секреции — гипофиз (hypophysis) (см. рис. 32, 9). На тонкой крыше промежуточного мозга развиваются два пузыревидных образования: передний называется теменным или париетальным органом, а задний — пинеальным органом или эпифизом (epyphysis) (см. рис. 32, 10, 11). У круглоротых оба образования выполняют роль светочувствительных органов; оба несут и секреторную функцию. У остальных позвоночных животных эпифиз функционирует только как железа внутренней секреции. Теменной (париентальный) орган сохраняется у некоторых рыб, земноводных и у части пресмыкающихся как светочувствительный орган; у остальных позвоночных исчезает.
Утолщенные стенки промежуточного мозга называют зрительными буграми (thalami optici) (см. рис. 32,13)\ выросты их стенок образуют зрительные нервы (п. optici) (II пара головных нервов), имеющие хиазму (перекрест): часть волокон нерва правой стороны уходит в левый нерв, а часть волокон слева переходит в правый нерв. Зрительные нервы иннервируют только сетчатку глаза. Позади хиазмы на дне промежуточного мозга образуется полый вырост — воронка (infundibulum), к передней стенке которого примыкает железа внутренней секреции — гипофиз (hypophysis) (см. рис. 32, 9). На тонкой крыше промежуточного мозга развиваются два пузыревидных образования: передний называется теменным или париетальным органом, а задний — пинеальным органом или эпифизом (epyphysis) (см. рис. 32, 10, 11). У круглоротых оба образования выполняют роль светочувствительных органов; оба несут и секреторную функцию. У остальных позвоночных животных эпифиз функционирует только как железа внутренней секреции. Теменной (париентальный) орган сохраняется у некоторых рыб, земноводных и у части пресмыкающихся как светочувствительный орган; у остальных позвоночных исчезает.
Крыша среднего мозга приподнята парными зрительными долями (lobi optici) и образует двухолмие (см. рис. 32, 23)\ у млекопитающих оно превращается в четверохолмие. Здесь в сером веществе заканчиваются волокна зрительного тракта. Начиная с двоякодышащих рыб и земноводных, значение среднего мозга как зрительного центра уменьшается. От него отходят две пары головных нервов, иннервирующих мышцы глаз: глазодвигательный нерв (n. oculomotorius) (III пара) и блоковый (п. trochlearis) (IV пара, рис. 34).
У круглоротых, части рыб и земноводных задний мозг имеет вид небольшой складки — мозжечка (см. рис. 32, 6), ограничивающей спереди крышу четвертого желудочка. У хрящевых рыб он разрастается, прикрывая заднюю часть среднего и переднюю часть продолговатого мозга. У пресмыкающихся и особенно у птиц и млекопитающих размеры мозжечка еще более возрастают, а поверхность его коры, образованной серым веществом, увеличивается благодаря образованию глубоких и сложных складок. Мозжечок — центр координации движений и равновесия; участвует он и в регуляции тонуса мышц и других физиологических процессов.
Продолговатый мозг (см. рис. 32, 7) имеет утолщенное дно и стенки, тогда как крыша его объемистой полости — четвертого желудочка (ромбовидной ямки) — образована лишь тонким эпителием и сосудистым сплетением мягкой мозговой оболочки. Значительную массу продолговатого мозга составляет белое вещество (в том числе и волокна, приходящие сюда из спинного мозга); его разрастания по дну мозга образуют так называемые пирамиды. Серое вещество располагается в верхней части дна и стенок продолговатого мозга; от него отходят V—X (у млекопитающих V—XII) пары головных нервов (рис. 34).
V пара — тройничный нерв (n. trigeminus) берет начало от боковой поверхности продолговатого мозга и тут же разделяется на три основные ветви: глазничную, верхнечелюстную и нижнечелюстную; иннервирует зубы и мускулатуру челюстной дуги, слизистую рта и кожу передней части головы. От дна мозга отходит тонкий отводящий нерв (п. abducens) (VI пара), иннервирующий наружную прямую мышцу глаза. За тройничным нервом несколькими корешками, сливающимися в крупный ганглий, отходит VII пара — лицевой нерв (n. facialis). Он распадается на мощную глазничную (идет вместе с одноименной ветвью V нерва), щечную, небную и подъязычную ветви; лицевой нерв иннервирует кожу головы, слизистую ротовой полости и мускулатуру подъязычной дуги. Далее начинается короткий слуховой нерв (n. acus- ticus), VIII пара, иннервирующий чувствующий эпителий внутреннего уха. Языкоглоточный нерв (n. glossopharingeus), IX пара, отходит позади слухового нерва и иннервирует слизистую глотки и мускулатуру первой жаберной дуги, а у наземных позвоночных — глотку, ее мускулатуру и вкусовые тельца языка. От заднебоковой поверхности продолговатого мозга многочисленными корешками, сливающимися сразу в крупный ганглий, отходит X пара — блуждающий нерв (n. vagus), разделяющийся затем на ряд ветвей: четыре жаберных (иннервируют слизистую глотки и мускулатуру II—V жаберных дуг), внутренностную (иннервирует сердце, кишечник, плавательный пузырь, легкие), боковую (иннервирует органы боковой линии; редуцируется у наземных позвоночных). XI пара — добавочный нерв (п. accessorius) образуется из задних корешков блуждающего нерва; хорошо обособлен только у млекопитающих и слабо различим у других амниот; иннервирует мускулатуру плечевого пояса. XII пара — подъязычный нерв (n. hypoglossus) хорошо выражен только у амниот, отходит от брюшной стенки задней части продолговатого мозга к мускулатуре языка и подъязычному аппарату.
I, II, VIII пары головных нервов — только чувствующие, III, IV, VI, XI и XII — только двигательные, V, VII, IX и X — смешанные (включают чувствующие и двигательные волокна).
Без отчетливых внешних границ продолговатый мозг переходит в спинной, лежащий в канале, образованном верхними дугами позвонков. Он имеет форму плоской ленты (круглоротые) или округлого тяжа; в нем имеется небольшая полость — невроцель. Вокруг нее концентрируется серое вещество (скопление нервных клеток с их дендритами и опорных клеток); у наземных позвоночных на поперечном разрезе спинного мозга оно по своему очертанию напоминает
Рис. 35. Поперечный разрез спинного мозга. Показано образование и ветвление спинного нерва:
/ — спинной корешок. 2 — брюшной корешок, 3 — спинальный ганглий,
4 — кожа, 5 — мускулы, в — спиниая ветвь, 7 — брюшная ветвь. 8 — ветвь к внутренним органам, 9 *- симпатический ганглий, 10 — кишечник,
// — спиниой мозг
крылья бабочки (рис. 35). В отличие от головного мозга в спинном мозге белое вещество (проводящие пути, образованные длинными отростками — аксонами нервных клеток с вкрапленными между ними опорными клетками) расположено снаружи. От боковой поверхности спинного мозга метамерно отходят спинномозговые нервы. От спинного выроста (рога) серого вещества отходит спинной корешок, состоящий из чувствующих волокон (передают нервные импульсы в головной мозг); на нем образуется расширение — спинномозговой узел (нервный ганглий). От брюшного выроста (рога) серого вещества отходит брюшной корешок, состоящий только из двигательных волокон (передают импульсы двигательной мускулатуре). Спинной и брюшной корешки сливаются в спинной нерв, выходящий между верхними дугами соседних позвонков и почти сразу же распадающийся на три ветви: спинную (иннервирует кожу и мышцы спины), брюшную (идет
в боковые и брюшные стенки тела) и внутренностную (соединяется с ганглием симпатической нервной системы) и иннервирует внутренние органы). Каждая^ из них включает как двигательные (эфферентные), так и чувствующие (афферентные) волокна.
В спинном мозге замыкаются рефлекторные дуги, независимо от головного мозга обеспечивающие элементарные безусловно рефлекторные акты, как, например, оборонительные. Нейроны, связывающие правые и левые стороны каждого сегмента спинного мозга, увеличивают эти возможности. Однако работа всего спинного мозга подчинена контролю головного мозга, осуществляемому через так называемые спинальные тракты — совокупности нервных волокон, по которым передается информация в головной мозг (восходящие) или распространяются импульсы, идущие от головного к сегментам спинного мозга (нисходящие пути). У бесчелюстных (круглоротых) восходящий путь не непрерывен, а состоит из коротких волокон Роон-Боардовских клеток, связывающих между собой соседние сегменты спинного мозга; этот путь кончается в продолговатом мозге. Нисходящий путь представлен идущими по всей длине спинного мозга волокнами Маутне- ровских клеток, имеющихся в среднем и продолговатом мозге (их считают близкими к гигантским волокнам мозга ланцетника), идущим от клеток Овсянникова-Роде. У хрящевых рыб к этим путям прибавляются два восходящих: спинно-бульбарный, кончающийся в продолговатом мозге, и спинно-мезэнцефалический, идущий в средний мозг. У костных рыб связи между головным и спинным мозгом усложняются путем образования двух новых путей — восходящего спинно-мозжечкового тракта и нисходящего вестибулярно-спинального. В других классах позвоночных усложнение этих связей усиливается и сопровождается все большим подчинением работы спинного мозга контролю головного. Следует обратить внимание на то, что в большинстве случаев новые пути связей образуются при сохранении старых систем, как некоторая надстройка над ними. Существование параллельных иерархически организованных каналов связи — важное условие эффективного управления работой всех систем организма, обеспечивающее ее изменение соответственно внешним условиям.
Такая адаптивность увеличивается и вегетативной нервной системой, возникающей в результате разрастания участков ганглионарных пластинок (см. рис. 31). При этом по бокам позвоночного столба образуются цепочки ганглиев вегетативной симпатической и парасимпатической [6] нервной системы. Длинные отростки клеток этих ганглиев направляются частично к внутренним органам (чувствующие и двигательные симпатические нервные волокна), частично входят в состав спинных корешков спинномозговых нервов. У двоякодышащих и костистых рыб и у всех наземных позвоночных ганглии вегетативной системы связаны друг с другом продольным нервным тяжом (truncus sympaticus). Соединительные ветви ганглиев обеспечивают связь центральной и вегетативной нервной системы. Чувствующие
и двигательные волокна нервов, отходящих от симпатических ганглиев, иннервируют внутренние органы, не подчиняющиеся волевому контролю (сердце, кишечник, почки, половые и эндокринные железы и др.). Показано, что вывод из строя вегетативной системы не ведет к гибели животного, но значительно снижает шансы выживания при отклонениях условий существования от нормы и при перенаселении.
Взаимосвязи центральной и вегетативной нервной системы создают возможность известной взаимозаменяемости нервных центров.
Гормональная система представлена разнообразными железами внутренней секреции. Ее сложность нарастает в эволюционном ряду позвоночных. При этом число и химизм участвующих в регуляции метаболизма гормонов в разных классах меняется мало, но существенно усложняются их функции: точнее — меняется реакция органов (тканей) на воздействие гормонов. Хорошим примером может служить пролактин из группы гонадотропных (ГТГ) гормонов гипофиза, который у млекопитающих стимулирует образование молока, функционирование желтого тела (в яичнике), влияет на придаточные железы половых органов самцов и оказывает общее воздействие на метаболизм при неспецифической стимуляции, например при стрессе. Пролактин у птиц стимулирует образование «зобного молочка» (у голубей), обусловливает поведение при насиживании яиц, вызывает образование «наседных пятен» и усиливает рост. У амфибий тот же пролактин регулирует проницаемость кожи для воды (тритоны), а у морских рыб способствует выживанию при осмотическом стрессе в пресной воде.
Деятельность отдельных звеньев гормональной системы взаимно координирована и находится под контролем центральной нервной системы; возникает сложный нейрогуморальный аппарат, координирующий все процессы в организме, включая особенности поведения и реакций на абиотические и биотические факторы внешней среды. Развитие нейрогуморального аппарата в ряду позвоночных животных приводит к становлению более сложных связей с окружающей средой и обеспечивает возрастание гомеостаза — внутренней устойчивости организма при меняющихся внешних воздействиях. Наконец, он участвует и в регуляции внутривидовых отношений и межвидовых связей в сообществе.
Органы чувств. Развитие и дифференцировка центральной нервной системы сопровождались возникновением и совершенствованием органов чувств — зрения, слуха и равновесия, обоняния, вкуса, восприятия движения воды (органы боковой линии водных позвоночных). Это улучшало возможности ориентации в пространстве, необходимость которой росла по мере увеличения подвижности животных.
Органы зрения позвоночных животных представлены парными глазами более или менее шаровидной формы, лежащими в орбитах — впадинах черепа. Слой плотной соединительной ткани или хряща образует наружную оболочку глаза — склеру (рис. 36, /). На передней поверхности глаза (перед зрачком) склера переходит в тонкую прозрачную роговицу, покрытую прозрачным эпителием. Изнутри
к склере прилегает сосудистая оболочка, обильно снабженная кровеносными сосудами1, за которой следует темная пигментная оболочка. На границе между склерой и роговицей края сосудистой и пигментной оболочек образуют кольцевую складку — радужину (рис. 36, 5), ограничивающую отверстие — зрачок, который может расширяться и сужаться благодаря присутствию в радужине радиальных и кольцевых мышечных волокон.
 Насыщение радужины пигментами обеспечивает различную окраску глаза. Позади радужины образуется кольцевой валик — ресничное тело, представляющее скопление мышечных волокон, прикрепляющихся к оболочке хрусталика. Ресничное тело участвует в аккомодации глаза, изменяя форму хрусталика, и, у части наземных позвоночных, перемещая его по отношению к сетчатке. За зрачком лежит круглое или сплюснутое в двояковыпуклую линзу прозрачное тело — хрусталик (рис. 36, 7).
Насыщение радужины пигментами обеспечивает различную окраску глаза. Позади радужины образуется кольцевой валик — ресничное тело, представляющее скопление мышечных волокон, прикрепляющихся к оболочке хрусталика. Ресничное тело участвует в аккомодации глаза, изменяя форму хрусталика, и, у части наземных позвоночных, перемещая его по отношению к сетчатке. За зрачком лежит круглое или сплюснутое в двояковыпуклую линзу прозрачное тело — хрусталик (рис. 36, 7).
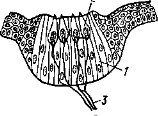
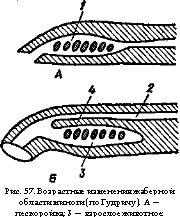
Внутренний слой стенки глаза — сетчатка — тесно примыкает к пигментной оболочке (рис. 36, 8). Она сложно устроена и состоит из светочувствительных (рецепторных), нервных и опорных клеток. К пигментному слою примыкает слой фоторецепторов (рис. 37, 2), наружные членики которых имеют вид палочек или колбочек (рис. 37, а и б) и содержат светочувствительный пигмент. Ядра светочувствительных клеток образуют наружный клеточный слой (рис. 37, в), а далее идет наружный сетчатый слой, где конечные разветвления чувствующих клеток контактируют с дендритами биполярных клеток. Отростки последних соприкасаются с дендритами ганглиозных клеток во внутреннем сетчатом слое (рис. 37, 5). Ганглиозные клетки (рис. 37, 6) дают длинные нервные волокна (аксоны), образующие внутренний слой сетчатки (рис. 37, 7) и далее составляющие зрительный нерв. В месте выхода зрительного нерва светочувствительный слой сетчатки редуцируется и этот участок называют «слепым пятном». В центре сетчатки обычно располагается область повышенной оптической чувствительности — желтое пятно; в его центре часто имеется углубление — ямка (см. рис. 36, 9), где плотность фоторецепторов и нервных клеток увеличена. Внутренняя полость глазного бокала (яблока) заполнена прозрачным студенистым стекловидным телом.
Движение глаза в орбите обеспечивается сокращением четырех прямых и двух косых мышц глаза, прикрепляющихся к стенкам глазницы
и склере. Вокруг глаза развивается складка кожи, которая у наземных позвоночных, начиная с амфибий, превращается в подвижное верхнее и нижнее веки. У многих позвоночных развивается еще тонкое полупрозрачное третье веко — мигательная перепонка. В области век развиваются железы, секрет которых смачивает роговицу, предохраняя ее от высыхания.

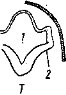
 Глаза позвоночных животных закладываются на ранних стадиях зародышевого развития. Одновременно с обособлением промежуточного мозга на его стенках образуются боковые выпячивания — глазные пузыри (рис. 38, 2). По мере роста зародыша глазные пузыри отодвигаются от мозга, их ножки удлиняются и сужаются, а наружная часть стенки каждого пузыря впячивается. Образуются два глазных бокала с двойными стенками, лежащие под кожей. Эктодерма над глазными бокалами образует утолщение, которое отшнуровывается от кожи, располагается у отверстия глазного бокала и превращается в хрусталик. Внутренняя, более тол- I стая, стенка глазного бокала преобразуется в
Глаза позвоночных животных закладываются на ранних стадиях зародышевого развития. Одновременно с обособлением промежуточного мозга на его стенках образуются боковые выпячивания — глазные пузыри (рис. 38, 2). По мере роста зародыша глазные пузыри отодвигаются от мозга, их ножки удлиняются и сужаются, а наружная часть стенки каждого пузыря впячивается. Образуются два глазных бокала с двойными стенками, лежащие под кожей. Эктодерма над глазными бокалами образует утолщение, которое отшнуровывается от кожи, располагается у отверстия глазного бокала и превращается в хрусталик. Внутренняя, более тол- I стая, стенка глазного бокала преобразуется в
* сетчатку, а более тонкая наружная — в пигмент
ный слой. Отверстие глазного бокала сужается. Из окружающей глазной бокал мезодермы формируются сосудистая оболочка, склера и роговица (рис. 38, 7—9); последнюю снаружи покрывает тонкий слой прозрачного эпителия — участок эктодермы.
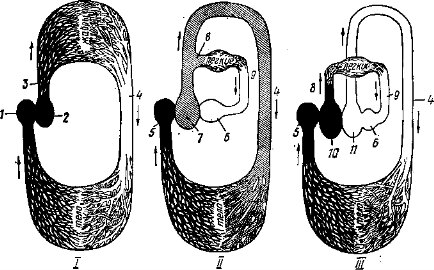 Схема строения глаза едина во всех классах позвоночных животных. Отличия сводятся к частным деталям строения и связанных с ним образований — век, глазных желез. Уже указывалось на существование у круглоротых добавочных светочувствительных органов: париетального (теменного) и пинеального (эпифиз). У части рыб, некоторых амфибий и пресмыкающихся имеется теменной орган, принимающий глазоподобное строение: утолщенная передняя стенка образует хрусталик, задняя стенка пигментирована и содержит светочувствительные клетки, палочковидные окончания которых (в отличие от настоящего глаза) направлены к внутренней полости теменного органа; под светочувствительными клетками в слое нервных волокон лежат гангоиозные клетки, отростки которых образуют нерв, идущий в крышу промежуточного мозга.
Схема строения глаза едина во всех классах позвоночных животных. Отличия сводятся к частным деталям строения и связанных с ним образований — век, глазных желез. Уже указывалось на существование у круглоротых добавочных светочувствительных органов: париетального (теменного) и пинеального (эпифиз). У части рыб, некоторых амфибий и пресмыкающихся имеется теменной орган, принимающий глазоподобное строение: утолщенная передняя стенка образует хрусталик, задняя стенка пигментирована и содержит светочувствительные клетки, палочковидные окончания которых (в отличие от настоящего глаза) направлены к внутренней полости теменного органа; под светочувствительными клетками в слое нервных волокон лежат гангоиозные клетки, отростки которых образуют нерв, идущий в крышу промежуточного мозга.
Органы слуха анатомически связаны с органами равновесия; у позвоночных животных они всегда парные (правое и левое «ухо»).
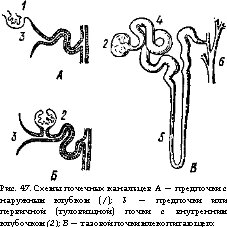
Рис. 38. Последовательные стадии развития глаза (I—IV — схема) (по Шмаль-
гаузену):
1 — промежуточный мозг, 2 — глазной пузырь, 3 — ножка глазного пузыря, 4 — хрусталик, 5 -» сетчатка, 6 — пигментный слой, 7 — сосудистая оболочка, 8 — склера, 9 — роговица
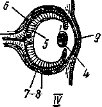
 alt="" />У водных позвоночных такой комплексный орган образован капсулами внутреннего уха, закладывающимися по бокам головы зародыша в виде парных утолщений эктодермы (рис. 39). Каждая из них превращается сначала в ямку, а затем в пузырек, сообщающийся с поверхностью каналом, и позднее отшнуровывающийся от эктодермы. Слуховой пузырек перехватом разделяется на два отдела: из верхнего формируется овальный мешочек и соединенные с ним три полукружных канала (рис. 39, 2—4), лежащие в трех взаимно перпендикулярных плоскостях, — орган равновесия (вестибулярный аппарат); нижний отдел превращается в круглый мешочек (рис. 39, 5), от которого отходит полый выступ — лагена, у высших позвоночных образующий спирально завитой канал (улитку) — орган слуха. С круглым мешоч-
alt="" />У водных позвоночных такой комплексный орган образован капсулами внутреннего уха, закладывающимися по бокам головы зародыша в виде парных утолщений эктодермы (рис. 39). Каждая из них превращается сначала в ямку, а затем в пузырек, сообщающийся с поверхностью каналом, и позднее отшнуровывающийся от эктодермы. Слуховой пузырек перехватом разделяется на два отдела: из верхнего формируется овальный мешочек и соединенные с ним три полукружных канала (рис. 39, 2—4), лежащие в трех взаимно перпендикулярных плоскостях, — орган равновесия (вестибулярный аппарат); нижний отдел превращается в круглый мешочек (рис. 39, 5), от которого отходит полый выступ — лагена, у высших позвоночных образующий спирально завитой канал (улитку) — орган слуха. С круглым мешоч-
Рис. 39. Схема развития внутреннего уха позвоночных животных. /—VII — последовательные стадии (по Шмальгаузену):
1 — овальный мешочек, 2—4 — полукружные каналы, б — круглый мешочек, S — эндолнм- фатнческий канал, 7 — полый выступ круглого мешочка — лагеиа, 8 — слуховые чувствительные поля, 9 — отолиты
ком связан длинный, обычно замкнутый, эндолимфатический канал — остаток соединения полости первичного слухового пузырька с внешней средой.
Эпителий, выстилающий перепончатый лабиринт, на некоторых участказКкруглого и овального мешочка и в полукружных каналах имеет чувствующие клетки, снабженные упругими волосками, вдающимися в просвет внутренней полости; основания грушевидных чувствующих клеток оплетаются разветвлениями конца слухового нерва. Внутренняя полость перепончатого лабиринта заполнена эндолимфой, в которой взвешены мелкие кристаллики извести — отоконии, иногда сливающиеся в крупные образования (отолиты костистых рыб). Всякое изменение положения головы вызывает перемещение эндолимфы и отокониев, раздражающих при этом чувствующие клетки. Перепончатый лабиринт, таким образом, функционирует как орган равновесия. Звуковые волны, вызывающие колебания эндолимфы и находящихся в ней включений, также вызывают раздражение чувствующих клеток. Однако слуховую функцию у рыб несут чувствующие поля полого выроста круглого мешочка. У наземных позвоночных его размеры заметно увеличиваются, и в образовавшейся таким путем улитке происходит усложнение воспринимающего аппарата за счет разрастания чувствующих клеток и добавочных мембран. Наряду с усложнением воспринимающего аппарата внутреннего уха у наземных позвоночных образуется среднее ухо, снабженное барабанной перепонкой и слуховыми косточками, а затем формируется и наружное ухо (хорошо развито у млекопитающих). Эти преобразования обеспечивают возможность слуха в воздушной среде, отличающейся более слабой звукопроводимостью. Часть лабиринта, функционирующая в качестве органа равновесия (овальный мешочек, полукружные
каналы), у всех позвоночных сохраняется примерно на одном уровне
развития; лишь у круглоротых он, видимо, вторично упрощен.
У водных позвоночных имеются специальные сейсмосенсорные кожные органы боковой линии. Они служат преимущественно для восприятия слабых колебанийтоков воды. У круглоротых и живущих в воде амфибий эти органы представляют собой лежащие на
поверхности или в мелких ямках утолщения небольших участков эктодермы, в
 которых среди опорных клеток лежат грушевидные чувствующие клетки (рис. 40): их выдающиеся над поверхностью части снабжены щетинками (жгутиками), а основания оплетены концевыми окончаниями боковой ветви блуждающего нерва. Эти органы располагаются в один — три продольных ряда на боковой поверхности тела и в несколько рядов, иногда в виде Рис. 40. Разрез органа бо- сложной сети — на голове. У рыб в толще к°в°й =гаамфибии (по кожи образуются каналы, открывающиеся
которых среди опорных клеток лежат грушевидные чувствующие клетки (рис. 40): их выдающиеся над поверхностью части снабжены щетинками (жгутиками), а основания оплетены концевыми окончаниями боковой ветви блуждающего нерва. Эти органы располагаются в один — три продольных ряда на боковой поверхности тела и в несколько рядов, иногда в виде Рис. 40. Разрез органа бо- сложной сети — на голове. У рыб в толще к°в°й =гаамфибии (по кожи образуются каналы, открывающиеся
наружу специальными отверстиями; у ко-
I — опорные клетки, 2 — чув- ^ г J
ствующие клетки, 3 — нерв СТИСТЫХ рыб ЭТИ ОТВерСТИЯ прОНИЗЫ
вают чешуи и покровные кости. На стенках каналов располагаются отдельные рецепторы боковой линии (рис.41,4), имеющие описанное строение.
 У рыб встречаются и отдельные органы боковой линии, лежащие на поверхности кожи. Улавливая органами боковой линии слабые токи воды, водные позвоночные могут ориентироваться в течениях, а воспринимая волны, идущие от плывущих животных или отраженные волны собственного перемещения, могут обнаруживать приближение хищника или добычи, плыть, не натыкаясь в темноте на подводные предметы. У наземных позвоночных органы боковой линии исчезают.
У рыб встречаются и отдельные органы боковой линии, лежащие на поверхности кожи. Улавливая органами боковой линии слабые токи воды, водные позвоночные могут ориентироваться в течениях, а воспринимая волны, идущие от плывущих животных или отраженные волны собственного перемещения, могут обнаруживать приближение хищника или добычи, плыть, не натыкаясь в темноте на подводные предметы. У наземных позвоночных органы боковой линии исчезают.
Орган обоняния развивается как утолщение эктодермы, которое затем погружается в кожу, образуя обонятельный мешок, открывающийся наружу отверстием — ноздрей. Эпителий обонятельного мешка состоит из опорных и удлиненных чувствующих клеток. Выходящие на поверхность эпителия концы чувствующих клеток несут пучок волосков, а проксимальные концы их соединяются с волокнами обонятельного нерва. У круглоротых парные вначале обонятельные мешки срастаются в один мешок, открывающийся наружу одной ноздрей. У остальных позвоночных органы обоняния парные. У некоторых рыб каждая ноздря разделяется кожистой перемычкой на два отверстия — через одно вода поступает в обонятельный мешок, а через
 второе выводится наружу. Образование складок на стенках обонятельного мешка увеличивает поверхность чувствующего эпителия. У двоякодышащих и кистеперых рыб полость обонятельного мешка через ноздрю открывается наружу, а через внутреннюю ноздрю или хоану сообщается с ротовой полостью.
второе выводится наружу. Образование складок на стенках обонятельного мешка увеличивает поверхность чувствующего эпителия. У двоякодышащих и кистеперых рыб полость обонятельного мешка через ноздрю открывается наружу, а через внутреннюю ноздрю или хоану сообщается с ротовой полостью.
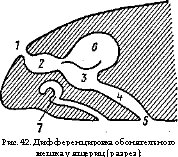
alt="" />Переход к воздушному дыханию и выход на сушу сопровождался превращением обонятельного тракта в дыхательно-обонятельный: воздух через наружные ноздри проходит в обонятельный мешок и через хоаны поступает в ротовую полость, гортань и легкие. В связи с этим уже у амфибий обонятельный мешок начинает подразделяться на дыхательный, или респираторный (выст
лан простым эпителием), и обонятельный, или ольфакторный (выстлан чувствующим эпителием) отделы. В обонятельном отделе обычно появляются складки, увеличивающие его поверхность. Еще отчетливее дифференцировка у настоящих наземных позвоночных (рис. 42). Особенно сложно строение обонятельного отдела у млекопитающих, у которых развиваются ажурные переплетения решетчатой кости, образующей костные обонятельные раковины с очень большой общей поверхностью, выстланной обонятельным эпителием.
Органами вкуса у позвоночных служат вкусовые почки, представляющие скопления чувствующих и опорных клеток. Основания чувствующих клеток оплетаются концевыми окончаниями нервов (лицевого, а у млекопитающих еще и языкоглоточного). Вкусовые почки у рыб есть в иищеводе, глотке и ротовой полости, а также на губах, усиках, иногда на плавниках и других участках тела, у остальных позвоночных они сосредоточены в ротовой области (преимущественно на языке и иногда в начальной части глотки).
Восприятие тактильных, химических, электрических (электромагнитных), температурных и других раздражений обеспечивается наличием свободных окончаний чувствующих нервов по всей поверхности кожи. В некоторых случаях они представляют специальные органы (электрорецепторы хрящевых рыб, см. ниже) или входят в состав системы органов боковой линии. Простое строение (свободные нервные окончания) имеют и так называемые интерорецепторы, собирающие информацию о состоянии внутренней среды и органов тела животного — химизме, температуре, осмотическом состоянии и других показателях. Информация от органов чувств (экстерорецеп- ция) и интерорецепторов поступает в соответственные отделы головного мозга, регулирующие физиологические параметры и поведение животного.
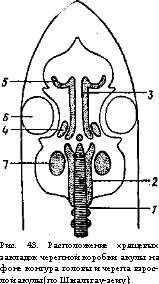
Скелет черепа. Развитие головного мозга и связанных с ним сложно устроенных органов чувств требовало образования защитных скелетных образований. Эту функцию выполняет мозговой или осевой череп. С другой стороны, переход к активному питанию подвижными пищевыми объектами должен был сопровождаться образованием органов активного захвата и переработки пищи. Это обеспечено формированием висцерального скелета, окружающего передний конец пищеварительной трубки; у водных позвоночных он служит и опорой жабр. Висцеральный и мозговой череп у всех позвоночных животных, кроме круглоротых, объединяется в единый комплекс — череп (cranium), подразделяемый на мозговой и висцеральный отделы.
Мозговой череп (neurocranium) закладывается под головным мозгом в виде двух-трех пар хрящей (рис. 43). По бокам переднего конца хорды образуются парахордалии (слившиеся зачатки верхних дуг передних позвонков), а впереди них — маленькие боковые хрящи (развиваются не всегда) и крупные трабекулы. Одновременно, но независимо от этих зачатков возникают хрящевые капсулы органов чувств: обонятельные, слуховые и склера глазных яблок. Парахордалии, трабекулы и боковые хрящи, разрастаясь, сливаются друг с другом, и начинают обрастать головной мозг с боков. К этой хряще
вой основе спереди прирастают обонятельные капсулы, а по бокам сзади — слуховые капсулы. Между ними на боковых поверхностях черепа образуются углубления — глазницы, в которых размещаются глаза. Такая стадия развития мозгового черепа, еще лишенного крыши, присуща круглоротым. У остальных позвоночных еще на личиночной или зародышевой стадии разросшийся хрящ образует н крышу черепа, в которой обычно остаются -небольшие отверстия —фонтанели, затянутые соединительнотканными перепонками. У низших костных рыб (хрящевых ганоидов) мозговой череп остается хрящевым, а поверх него образуется панцирь из покровных (кожных) костей. У остальных групп костных рыб в первичном хрящевом мозговом черепе возникают окостенения, завершающиеся образованием так называемых первичных, или хрящевых, костей. Одновременно покровные кости погружаются под кожу и вступают в соединение с хрящевыми костями. По внешнему виду и гистологическому строению покровные и хрящевые кости очень похожи; различить их можно лишь проследив развитие. У костистых рыб мозговой череп состоит из большого числа хрящевых и покровных костей; хрящ сохраняется только в области обонятельных и слуховых капсул и глазниц. У современных амфибий в мозговом черепе остаются значительные участки хряща и развивается относительно небольшое число хрящевых и покровных костей. У рептилий, птиц и млекопитающих мозговой череп во взрослом состоянии образован лишь костями (хрящевыми и покровными).
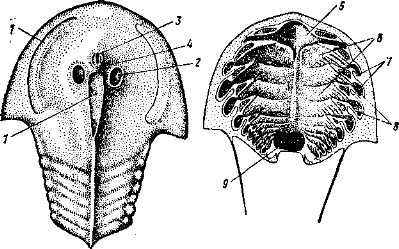
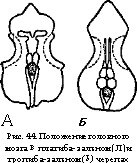
 Различают два основных типа мозгового (осевого) черепа: 1) плати- базальный — с широким основанием; между его глазницами расположена мозговая полость черепа (рис. 44, А) — свойствен многим группам рыб, амфибий, части рептилий; 2) тропибазальный — с узким основанием; стенки глазниц сближены и разделены лишь тонкой меж- глазничной перегородкой; мозговая полость расположена позади глазниц (рис. 44, Б).
Различают два основных типа мозгового (осевого) черепа: 1) плати- базальный — с широким основанием; между его глазницами расположена мозговая полость черепа (рис. 44, А) — свойствен многим группам рыб, амфибий, части рептилий; 2) тропибазальный — с узким основанием; стенки глазниц сближены и разделены лишь тонкой меж- глазничной перегородкой; мозговая полость расположена позади глазниц (рис. 44, Б).
Висцеральный отдел черепа (splanchnocranium) развивается независимо от мозгового черепа в виде висцеральных (жаберных) дуг, лежащих в перегородках между жаберными щелями. У круглоротых эти хрящевые зачатки преобразуются в сложную хрящевую жаберную коробку (окружает область жаберных мешков), в околосердечный хрящ и в хрящи, поддерживающие мускулатуру языка и ротовой воронки. У водных челюстноротых позвоночных животных рудименты
первых двух жаберных дуг образуют губные хрящи (развиты у акуловых рыб). Третья висцеральная дуга, которую называют челюстной, расчленяется на два отдела, образуя собственно челюсти: верхний элемент называют небно-квадратным хрящом (palatoquad- ratum), нижний — меккелевым хрящом (carti- lago meckeli). Эти элементы выполняют функцию челюстей только у низших рыб. У высших рыб они окостеневают и лишь частично участвуют в удержании добычи; основная функция захвата добычи переходит к вторичным челюстям, т. е. к покровным, по происхождению, костям: верхнечелюстной (maxil- lare) и предчелюстной (praemaxillare) — в верхней челюсти; зубной (dentale) — в нижней челюсти. Вторичные челюсти связаны с костями, возникшими в результате окостенения первичных хрящевых челюстей.
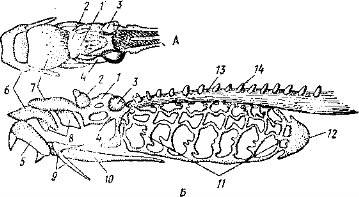 Следующая висцеральная дуга, подъязычная, обычно состоит из двух крупных парных хрящевых элементов: гиомандибулярного, или подвеска (hyomandibulare), и гиоида, или подъязычного (hyoideum). Гиоиды правой и левой сторон соединяются друг с другом при помощи небольшого непарного элемента — копулы (copulae); у высших рыб подъязычная дуга окостеневает. Остальные дуги служат опорой жабр (жаберные дуги) и обычно состоят из четырех подвижно сочлененных друг с другом парных элементов; правая и левая половина каждой дуги соединяется друг с другом на брюшной стороне при помощи непарного элемента. У примитивных рыб число дуг может достигать 7 пар, у большинства хрящевых рыб — 5 пар хрящевых жаберных дуг, у костистых рыб часто их 4 пары (V-рудиментарна) и они окостеневают. За счет покровных костей у костных рыб формируется скелет жаберной крышки, тоже относящийся к висцеральному черепу.
Следующая висцеральная дуга, подъязычная, обычно состоит из двух крупных парных хрящевых элементов: гиомандибулярного, или подвеска (hyomandibulare), и гиоида, или подъязычного (hyoideum). Гиоиды правой и левой сторон соединяются друг с другом при помощи небольшого непарного элемента — копулы (copulae); у высших рыб подъязычная дуга окостеневает. Остальные дуги служат опорой жабр (жаберные дуги) и обычно состоят из четырех подвижно сочлененных друг с другом парных элементов; правая и левая половина каждой дуги соединяется друг с другом на брюшной стороне при помощи непарного элемента. У примитивных рыб число дуг может достигать 7 пар, у большинства хрящевых рыб — 5 пар хрящевых жаберных дуг, у костистых рыб часто их 4 пары (V-рудиментарна) и они окостеневают. За счет покровных костей у костных рыб формируется скелет жаберной крышки, тоже относящийся к висцеральному черепу.
Различают несколько типов прикрепления челюстного аппарата к мозговому черепу. 1. Протостилия — челюстная и подъязычная дуги независимо друг от друга связками подвешивались к мозговому черепу. Этот гипотетический исходный тип черепа, вероятно, был свойствен примитивным челюстноротым. 2. Гиостилия — верхний конец гиомандибулярного отдела подъязычной дуги прикрепляется к слуховому отделу мозгового черепа и служит подвеском для челюстной дуги (многие группы рыб, в том числе большинство хрящевых и все костистые рыбы). 3. Амфистилия — верхний элемент челюстной дуги соединяется с мозговым черепом при помощи одного-двух специальных отростков и, кроме этого, как и при гиостилии, задние концы обоих челюстных элементов прочно связаны с нижним концом гиомандибулярного элемента (некоторые примитивные и современные акулы, костные ганоиды). 4. Аутостилия — верхний элемент челюстной дуги соединяется или срастается с мозговым черепом, гиомандибуляр- ный элемент подъязычной дуги в прикреплении челюстей не участвует, и подъязычная дуга в большей или меньшей степени подвергается
редукции (цельноголовые и двоякодышащие рыбы, земноводные и все другие наземные позвоночные).
С переходом к воздушному дыханию висцеральный скелет видоизменяется. В ряду земноводные — млекопитающие происходит усиление вторичных челюстей, а развитие аутостилии приводит к редукции подъязычной дуги: нижний ее элемент вместе с редуцирующими жаберными дугами участвует в образовании подъязычного аппарата и скелета гортани, а верхний элемент — гиомандибулярный, или подвесок, — превращается в косточку среднего уха — стремечко. Скелет жаберной крышки полностью редуцируется.
Пищеварительная система существенно усложняется. У древних бесчелюстных рот сосущий, у современных круглоротых развивается присасывательная воронка и аппарат всасывания крови и лизирован- ных тканей жертвы (особенно у миксии). У челюстноротых образование вооруженных зубами челюстей обеспечило возможность захвата, удержания, а у некоторых и механической обработки пищи. Дифференцируется жевательная мускулатура, управляющая движением челюстей. Преобразования зубов и челюстного аппарата (включая жевательную мускулатуру) сопровождались расширением спектра питания. Отчетливо проявляется процесс дифференцировки и удлинения пищеварительной трубки, разделяющейся на ротовую полость, глотку, пищевод, желудок и несколько отделов кишечника.
На дне ротовой полости имеется язык с собственной мускулатурой и скелетом (подъязычный аппарат); у рыб он представляет лишь складку слизистой оболочки. В слизистой языка и всей ротовой полости рассеяны вкусовые и осязательные тельца. Начиная с амфибий в ротовой полости появляются слюнные железы. Их секрет — слюна — увлажняет слизистую рта, предохраняя ее от высыхания, и смачивает пищу, облегчая ее проглатывание. У змей отдельные слюнные железы преобразуются в ядовитые железы. Липкая слюна некоторых амфибий, рептилий, птиц и млекопитающих обеспечивает прилипание к языку мелкой подвижной добычи. У млекопитающих слюна содержит и пищеварительный фермент птиалин (амилазу), расщепляющий углеводы (крахмал).
Глотка у водных позвоночных животных пронизана жаберными щелями; у наземных позвоночных с глоткой связаны легкие. За глоткой идет пищевод — тонкая, растяжимая трубка, без резкой границы переходящая в желудок. Последний представляет расширение кишечной трубки, с более или менее развитой мускулатурой стенок; это обеспечивает перемешивание, а у некоторых групп, особенно птиц, и перетирание пищи. В слизистой желудка расположены трубчатые пищеварительные железы, выделяющие имеющий кислую реакцию желудочный сок. В его составе преобладает пепсин, но входят и ферменты химозин и липаза. Размеры, форма и детали строения желудка во всех классах позвоночных варьируют в очень широких пределах, отражая пищевую специализацию отдельных групп.
Кишечник обычно дифференцируется на три-четыре отдела: тонкая кишка (в ее переднюю часть, называемую двенадцатиперстной кишкой, открываются протоки печени и поджелудочной железы),
толстая кишка и задняя (или прямая) кишка, открывающаяся в клоаку или самостоятельным анальным отверстием; на границе между тонкой и толстой кишками развивается слепая кишка (в зачаточном состоянии она есть у пресмыкающихся, обычно слабо развита у птиц и хорошо — у большинства млекопитающих). Слизистая кишечника содержит слизистые и пищеварительные железы, выделяющие слизистый секрет муцин (предотвращает самопереваривание тканей стенок кишечной трубки), комплекс пищеварительных ферментов и гормоны, регулирующие процесс переваривания. Хорошо развита мускульная оболочка, осуществляющая перистальтику кишечника и движение пищевых масс. В кишечник поступают образующаяся в печени желчь и ферменты поджелудочной железы. В передних отделах кишечника идет химическая переработка пищи (переваривание) и ее всасывание; в задней кишке происходит преимущественно всасывание воды и формирование каловых масс. Внутренняя поверхность кишечника увеличивается за счет развития складки (спиральный клапан круглоротых и низших рыб) либо удлинением кишечника, образующего петли. Помимо этого, увеличение всасывательной поверхности достигается развитием мелкой складчатости слизистой или образованием на ней многочисленных мелких сосочков — ворсинок. Особенно многочисленны и хорошо развиты ворсинки в кишках птиц и млекопитающих. Часть слизистой оболочки ротовой и клоакальной полостей образуется из эктодермы, а слизистая всего остального кишечного тракта — из первичной кишки, или энтодермы.
Печень и поджелудочная железа хорошо развиты у всех позвоночных животных. В эмбриогенезе печень развивается как слепой вырост, отшнуровывающийся от кишечника и превращающийся в компактную железу. В печени вырабатывается желчь, которая по желчному протоку (часто предварительно накапливаясь в желчном пузыре) поступает в начальную часть тонкой кишки. Желчь эмульгирует жиры и активизирует расщепляющую жиры липазу — фермент, выделяемый поджелудочной железой; благодаря своей щелочной реакции желчь нейтрализует кислый желудочный сок и возбуждает перистальтику кишечника. Кроме того, печень — важный кровеочистительный орган: в ней нейтрализуются поступающие в кровь вредные для тканей продукты распада. Наконец, в печени синтезируется гликоген (животный крахмал) и некоторые витамины (К)- Запасы гликогена служат энергетическим резервом организма, а витамин К играет важную роль в механизмах свертывания крови.
Поджелудочная железа развивается из нескольких мелких выпячиваний кишки в области печеночного выроста и превращается в компактную или дольчатую железу, лежащую на брыжейке начальной части тонкой кишки. Поджелудочная железа выделяет комплекс пищеварительных ферментов, среди них — трипсин (переваривает белки), амилаза (расщепляет углеводы — крахмал и др.) и липаза (переваривает жиры). В ткани поджелудочной железы обособляются островки Лангерганса, представляющие орган внутренней секреции, гормоны которого (инсулин и глюкагон) регулируют углеводный обмен.
У высших рыб (хрящевые и костные ганоиды, костистые рыбы) как вырост спинной части начала кишечника образуется плавательный пузырь, выполняющий функцию гидростатического органа и барорецептора, а иногда и добавочного органа дыхания.
 Дыхательная система. У первичноводных позвоночных животных (бесчелюстные и рыбы), как и у низших хордовых, образуются жаберные щели, сообщающие полость глотки с внешней средой.
Дыхательная система. У первичноводных позвоночных животных (бесчелюстные и рыбы), как и у низших хордовых, образуются жаберные щели, сообщающие полость глотки с внешней средой.
 У бесчелюстных (круглоротых) в жаберных щелях развиваются
У бесчелюстных (круглоротых) в жаберных щелях развиваются
складчатые стенки. У водных челюстноротых на перегородках между жаберными щелями развиваются складки слизистой оболочки — жаберные лепестки; их совокупность составляет жабры. К жаберным лепесткам кровь приносится артериями, а в них развивается густая сеть капилляров. Обмен газами в жабрах позвоночных построен по типу так называемых «противоточных систем»: при встречном движении кровь контактирует со все более богатой кислородом водой, что обеспечивает ее эффективное насыщение (рис. 45). Увеличение поверхности поглощения кислорода благодаря образованию жабр сопровождалось уменьшением числа жаберных щелей у позвоночных по сравнению с низшими хордовыми. У цельноголовых (из хрящевых рыб) намечается редукция межжаберных перегородок и образуется кожистая (не имеющая костей) жаберная крышка, прикрывающая снаружи область жаберных щелей. У костных рыб в жаберной крышке возникает костный скелет, а межжаберные перегородки редуцируются, что способствует более интенсивному омыва- нию водой жаберных листков. Основная функция жабр — газообмен: поглощение кислорода и выделение углекислоты. Наряду с этим жабры рыб выполняют и роль добавочных органов водного и солевого обмена (поглощают и выделяют воду и ионы солей, особенно хлористого натрия), способствуя поддержанию устойчивого осмотического давления крови и тканевых жидкостей, выделяют в значительных количествах аммиак и мочевину. В качестве добавочных органов дыхания у отдельных групп рыб функционируют кожа, плавательный пузырь и специализированные участки кишечной трубки (см. ниже).
У многоперовых и двоякодышащих рыб в виде парного выпячивания задней брюшной части глотки развиваются полые образования, внешне напоминающие плавательный пузырь, коротким каналом открывающиеся на брюшной стенке начальной части пищевода. Они функционируют как легкие. У личинок земноводных образуются жаберные щели и внутренние и наружные жабры, редуцирующие во время метаморфоза и заменяемые легкими, возникающими как парные выпячивания брюшной части глотки в области последней жаберной щели.
У первично наземных позвоночных — амниот (пресмыкающихся, птиц и млекопитающих) на относительно ранних стадиях зародышевого развития возникают зачатки жаберных щелей, вскоре исчезающие. На брюшной стороне глотки обособляется непарный желоб, у конца которого возникают парные выросты, превращающиеся в легкие; желоб превращается в трахею (воздуховод). Внутренняя поверхность легких резко увеличивается благодаря развитию сложной сети перекладин и складок, разделяющих полость легкого на мелкие ячейки. Интенсификации дыхания способствует не только рост внутренней поверхности легких, но и образование грудной клетки, обеспечивающей интенсивное вентилирование легких. В зародышевом периоде у высших позвоночных (амниот) в качестве основного органа дыхания функционирует зародышевый мочевой пузырь — аллантоис. С момента вылупления (рождения) функция дыхания переходит к легким.
Кровеносная система. Как и у бесчерепных, кровеносная система позвоночных замкнутая: кровь циркулирует по системе кровеносных сосудов, стенки которых имеют гладкие мускульные волокна и тонкую внутреннюю эндотелиальную оболочку; через такую систему биологических мембран обеспечивается активный обмен веществами между кровью и тканевой жидкостью. Замыкание кровеносной системы и появление эндотелиальной выстилки стенок кровеносных сосудов, наметившееся еще у бесчерепных, привело к появлению в организме трех сред: внутриклеточной, внутритканевой с межклеточной жидкостью— лимфой, и кровяного русла с кровью. Такая организация внутренней среды тела позвоночных обеспечивает ее устойчивость, необходимую для протекания биохимических процессов в подвижном организме, быстро меняющем как места пребывания (внешние условия), так и свое внутреннее состояние. Замыкание кровеносной системы, анатомически кажущееся малозначительным событием в эволюции, в действительности имело большое приспособительное значение; с ним связано оформление особой лимфатической системы (см. ниже).
Важное эволюционное приобретение позвоночных — образование сердца — специального органа» обеспечивающего ток крови по сосудам тела. Сердце возникло как расширение брюшной аорты. Его стенки образованы поперечнополосатой мускулатурой, тогда как в стенках кровеносных сосудов имеются лишь гладкие мышечные волокна. Сердце может быть двухкамерным (круглоротые, рыбы), т. е. состоять их одного предсердия (atrium) и одного желудочка (ventriculus), трехкамерным (земноводные, пресмыкающиеся) — из двух предсердий и одного желудочка, и четырехкамерным (птицы, млекопитающие) — из двух предсердий и двух желудочков. Дополнительные отделы сердца — венозная пазуха и артериальный конус; степень их развития варьирует в разных классах. В зависимости от направления движения крови различают два типа кровеносных сосудов: по более толстостенным артериям (arteriae) кровь течет от сердца (из желудочка), по венам (venae) она двигается к сердцу, попадая в венозную пазуху и предсердие. Самые мелкие разветвления артерий распадаются на мельчайшие кровеносные сосуды — капилляры, затем соединяющиеся в вены. В немногих случаях вены также могут распадаться
Рис. 46. Схема кровообращения позвоночных животных (по Бобринскому и Матвееву, 1966), I — первичноводное; II — земноводное; III — высшее наземное позвоночное; черным цветом показана венозная кровь, белым — артериальная, штриховкой — смешанная:
— предсердие, 2 — желудочек, 3 — брюшная аорта, 4 — спинная аорта, 5 — правое предсердие, 6 — левое предсердие, 7 — общий желудочек, 8 — легочная артерия, 9 — легочная вена, 10 — правый желудочек, 11 — левый желудочек. Стрелки указывают направление тока
крови
на капилляры, вновь объединяющиеся в вены. Тогда говорят об образовании воротной системы (воротная система печени, почек, гипофиза и др.).
У водных позвоночных (круглоротые, рыбы) только один круг кровообращения (рис. 46, /): из желудочка двухкамерного сердца кровь идет в жабры, где насыщается кислородом; по ответвлениям спинной аорты кровь разносится по всему телу, отдавая питательные вещества и кислород и насыщаясь продуктами распада; по венам венозная кровь возвращается в сердце (в предсердие). У земноводных и пресмыкающихся возникают два круга кровообращения (намечаются у двоякодышащих рыб): малый (легочный) и большой, четко не разделенные друг от друга, так как при трехкамерном сердце кровь из предсердий поступает в единый (хотя иногда перегородками частично разделенный) желудочек. Малый круг начинается в желудочке, включает легкие и завершается в левом предсердии. Большой круг кровообращения: от желудочка по сосудам всего тела в правое предсердие (рис. 46, II). В левое предсердие попадает артериальная кровь из легких, в правое ¦— венозная кровь со всего тела и — у земноводных — примесь артериальной крови, окислившейся в капиллярах кожи и ротовой полости. При одновременном сокращении предсердий кровь поступает в желудочек; развитие в его полости неполных перегородок и карманов уменьшает, но не предотвращает перемешивание крови.
Разделение потоков крови по ее составу при выходе из желудочка (наиболее богатая кислородом — в голову, наиболее бедная — в органы дыхания) обеспечивается расположением перегородок, работой клапанов желудочка и местом отхождения от него основных артериальных стволов. Этот тип кровеносной системы можно рассматривать как переходный между типично водным и типично наземным типами кровеносной системы.
У птиц и млекопитающих (рис. 46, ///) образуются полностью разделенные два круга кровообращения, что обеспечивается четы- рехкамерностыо сердца и полной изолированностью его венозной правой части от левой, артериальной. Малый круг кровообращения: правый желудочек — легочные артерии— легкие —легочные вены — левое предсердие. Большой круг: левый желудочек — аорта — артерии к голове, конечностям, внутренним органам — вены, впадающие в правое предсердие (рис. 46, III). С преобразованиями сердца, впадающих и отходящих от него кровеносных стволов идет перестройка периферической системы кровеносных сосудов.
В ряду позвоночных животных (круглоротые — млекопитающие) интенсифицируется кроветворение. У круглоротых форменные элементы крови (эритроциты, лимфоциты) образуются в жаберных лепестках, лимфоидной ткани кишечника и в почках. У рыб образование эритроцитов идет в селезенке, почках, отчасти в кровяном русле и стенке кишечника, лейкоцитов — в селезенке, ткани почек и других участках тела. У земноводных эритроциты образуются в селезенке, в костном мозге впервые образовавшихся крупных трубчатых костей и кровяном русле; лейкоциты — в печени и в почках. У рептилий, птиц и млекопитающих эритроциты образуются главным образом в костном мозге и менее в селезенке и в кровяном русле, а лимфоциты — в лимфоидных участках селезенки, печени, костного мозга и в лимфатических узлах (особенно у млекопитающих); утрачивая роль основного органа кроветворения, селезенка становится основным депо крови.
В ряду позвоночных животных увеличивается число эритроцитов, количество гемоглобина и объем крови (табл. 2). Одновременно повышается кровяное давление и растет буферность крови: она обеспечивается щелочным резервом (бикарбонаты — углекислота). Высокое содержание белков, сахаров и других веществ в плазме крови обусловливает ее энергетические и защитные свойства (иммунитет). Широкие колебания всех этих показателей в пределах каждого класса определяются экологическими особенностями отдельных видов (степенью их подвижности, богатством кислорода в среде и др.). Депонирование крови в ряде органов (печень, селезенка, кожа и др.) позволяет быстро восстановить потери при кровотечениях и увеличить объем циркулирующей крови при усилении движений.
Лимфатическая система. При замкнутой кровеносной системе кровь не является жидкой средой, окружающей клетки. Эту роль выполняет тканевая (межклеточная) жидкость — лимфа. У позвоночных обособляется лимфатическая система, включающая разного диаметра лимфатические сосуды и полости. Крупные сосуды имеют
соединительнотканные стенки с мышечными волокнами; их внутренняя оболочка образует складки — клапаны, допускающие ток лимфы лишь в одном направлении. Мелкие сосуды (лимфатические капилляры) со стенками из однослойного эпителия открываются непосредственно в межклеточные пространства. Собираемая капиллярами лимфа изливается в вены. Ток лимфы обеспечивается сжатием лимфатических сосудов окружающими мышцами и органами, а также пульсацией расширений этих сосудов — лимфатических сердец. Способствует току лимфы и присасывающее действие сердца. По ходу лимфатических сосудов расположены лимфатические железы — особенно выраженные у млекопитающих; в них образуются белые кровяные тельца — лимфоциты и происходит фагоцитоз проникших в организм болезнетворных агентов.
Таблица 2. Изменения количества крови и ее кислородной емкости в разных классах позвоночных животных (по П. А. Коржуеву, 1964, с изменен.)
|
Класс |
Количество крови в % от массы тела |
Количество гемоглобина |
Число эритроцитов в I мм3 крови в мли. шт. |
|
|
в г на 1 кг массы тела |
в г% в крови |
|||
|
Круглоротые |
4,0—5,0 |
•gt; |
3,0—8,0 |
0,13—0,17 |
|
Рыбы |
1.1—73 |
0,5—^-3,4 |
0,8—17,4 |
0,09—3,9 |
|
Земноводные |
12—7,2 |
0,2—4,8 |
1,9-10,0 |
0,02—0,7 |
|
Пресмыкающиеся |
3,9—8,3 |
2,1—4,9 |
4,0—11,0 |
0,5—1,6 |
|
Птицы |
6,5—15,6 |
4,6—17,8 |
|7,0-20,0 |
1,6—6,0 |
|
Млекопитающие |
4,8—Щ0 |
4,9-21,1 |
8,0-23,0 |
3,0—25,0 |
Выделительная система. С возрастанием уровня обмена веществ в ряду позвоночных животных шло совершенствование органов выделения; ими служат парные почки (renes). У зародышей (личинок) позвоночных образуется головная почка или предпочка (pronephros). Она представляет собрание нефридиальных канальцев,, открывающихся мерцательными воронками (нефростомамн) в полость тела, а другими концами — в собирательные канальцы, впадающие в выводной проток пронефроса. Около части нефростомов в стенках полости тела возникают грушевидные выросты из клубочков артериальных капилляров (рис. 47, А — 1). Они выделяют плазму крови, содержащую как продукты распада, так и полезные вещества. Образующиеся в головной части нефридиальных канальцев капсулообразные расширения охватывают такие клубочки сосудов, создавая боуменовы капсулы. Так образуется основной элемент почки позвоночных — мальпигиево тельце (рис. 47, 2). Продукты распада попадают в почечные канальцы пронефроса из целомической жидкости через нефро- стомы, а из крови — путем фильтрации через сосудистые клубочки. Черты строения пронефрической почки сохранились в дефинитивной мезонефрической почке взрослых миног.
Еще в зародышевом (личиночном) состоянии позади про- нефроса образуется первичная, или туловищная, почка (mesonephros). Часть почечных канальцев мезонефроса также имеют нефростомы (воронки) и мальпигиевы тельца, тогда как большинство их теряет воронки, имея только хорошо развитые мальпигиевы тельца. Возвращение в кровяное русло содержащихся в фильтрате мальпигиевых телец ценных веществ (воды, сахаров, витаминов и др.) производится в выводных канальцах.
 alt="" />Ко времени образования мезонефроса проток предпочки (пронефроса) у хрящевых рыб расщепляется на два канала: вольфов и мюллеров; у других позвоночных мюллеров канал возникает как новообразование. В вольфов канал открываются пртоки мезонефроса. У самцов анам- ний отходящие от семенника семявыносящие канальцы впадают в переднюю (у двоякодышащих рыб в заднюю) часть мезонефрической почки, которая теряет выделительную функцию, превращаясь фактически в придаток семенника. Поэтому у самцов анамний вольфов канал выполняет функцию и мочеточника, и семяпровода (иногда он вновь расщепляется или образует внутри почки длинные выросты,* благодаря чему половой и мочевой тракты разобщаются); пронефрос и мюллеров канал у них редуцируются. У самок вольфов канал выполняет только функцию мочеточника; тогда как мюллеров канал становится яйцеводом (рис. 48, II, III); при этом один из нефро- стомов пронефроса превращается в воронку яйцевода. Созревшая в яичнике яйцеклетка разрывает оболочку фолликула, выпадает в полость тела и через воронку попадает в мюллеров канал — яйцевод; нижняя часть яйцевода часто образует расширение — матку. Мюллеров и вольфов каналы открываются в клоаку. В брюшной части клоаки обычно образуется тонкостенное выпячивание — мочевой пузырь (рис. 48, 9). У части костистых рыб парные вольфовы каналы служат только мочеточником и у самцов и у самок; они впадают в мочевой пузырь. У обоих полов обособляются короткие самостоятельные половые протоки, открывающиеся наружу.
alt="" />Ко времени образования мезонефроса проток предпочки (пронефроса) у хрящевых рыб расщепляется на два канала: вольфов и мюллеров; у других позвоночных мюллеров канал возникает как новообразование. В вольфов канал открываются пртоки мезонефроса. У самцов анам- ний отходящие от семенника семявыносящие канальцы впадают в переднюю (у двоякодышащих рыб в заднюю) часть мезонефрической почки, которая теряет выделительную функцию, превращаясь фактически в придаток семенника. Поэтому у самцов анамний вольфов канал выполняет функцию и мочеточника, и семяпровода (иногда он вновь расщепляется или образует внутри почки длинные выросты,* благодаря чему половой и мочевой тракты разобщаются); пронефрос и мюллеров канал у них редуцируются. У самок вольфов канал выполняет только функцию мочеточника; тогда как мюллеров канал становится яйцеводом (рис. 48, II, III); при этом один из нефро- стомов пронефроса превращается в воронку яйцевода. Созревшая в яичнике яйцеклетка разрывает оболочку фолликула, выпадает в полость тела и через воронку попадает в мюллеров канал — яйцевод; нижняя часть яйцевода часто образует расширение — матку. Мюллеров и вольфов каналы открываются в клоаку. В брюшной части клоаки обычно образуется тонкостенное выпячивание — мочевой пузырь (рис. 48, 9). У части костистых рыб парные вольфовы каналы служат только мочеточником и у самцов и у самок; они впадают в мочевой пузырь. У обоих полов обособляются короткие самостоятельные половые протоки, открывающиеся наружу.
У зародышей первично наземных позвоночных (амниот) образуется пронефрос, потом закладывается мезонефрос и возникают вольфов и мюллеров каналы. Но во второй половине зародышевого развития в тазовой области образуются канальцы вторичной, или тазовой, почки (metanephros); они удлиненны и извиты, не имеют воронок и заканчи-
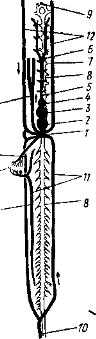
Рис. 48. Схема взаимоотношений половой н выделительной систем позвоночных животных (по Шмальгаузену, с изменен.) I — нейтральное зародышевое состояние; II — самка анамний; III — самец анамний; IV — зародыш амниот; V — самка амниот; VI — самец амниот (пунктиром обозначены редуцирующиеся отделы): — пронефрос, 2 — мезонефрос, 3 — метанефрос, 4 — канал проиефроса, 5 — мюллеров канал, выполняющий у самок функцию яйцевода, 6 — вольфов канал, выполняющий у самцов анамннй функцию мочеточника и семяпровода, у самцов амннот — только семяпровода, 7 — матка, 8 — мочеточник метанефрнческой почкн, 9 — мочевой пузырь, 10 — клоака, — мочеполовой сннус, 12 — половой член у самцов нлн клнтор у самок, 13 — половая железа, 14 — придаток семенника — остаток мезонефрической почки, 15 — семенник, 16 —
яичннк, 17 — воронка яйцевода, 18 — задняя кншка
ваются мальпигиевыми тельцами (рис. 48, IV—VI). При формировании метанефрнческой почки задний конец вольфова канала дает боковое выпячивание, врастающее в ткань метанефроса; в него прорываются канальцы метанефроса и он превращается в мочеточник (рис. 48, 8). Вольфов канал у самок редуцируется, а остатки мезонефроса превращаются в лимфоидную ткань; мюллеров канал сохраняется и функционирует как яйцевод. У самцов передняя часть мезонефроса, в которую открываются семявыносящие каналы семенников, превращается в придаток семенника (epididymis), а вольфов канал продолжает выполнять функцию семяпровода (рис. 48, 14, 6). В связи с клоакой формируются совокупительные органы. У млекопитающих клоака исчезает и формируется самостоятельное мочеполовое и анальное отверстия.
Половая система. Позвоночные животные, как правило, раздельнополы. Половые железы обычно парные. Яичники (ovarii) имеют более или менее заметное зернистое строение. Семенники (testiculi) отличаются гладкой поверхностью.
Для анамний характерно наружное оплодотворение, но у хрящевых и некоторых костных рыб, хвостатых и безногих земноводных возникает внутреннее оплодотворение. Яйца анамний способны развиваться только в водной (или, в редких случаях, в очень влажной) среде. Лишь у немногих групп появляется яйцеживорождение (задержка развивающегося яйца в нижних частях яйцеводов), настоя
щее живорождение (когда устанавливается обмен между развивающимся зародышем и материнским организмом, например у некоторых акуловых рыб) или развитие яйца идет в специальных наружных складках кожи (игла рыба, сумчатые квакши, пипа и др.). Яйца имеют наружную белковую оболочку, обеспечивающую защиту от механических и химических повреждений (иногда приобретающую большую прочность и водонепроницаемость — у миксин и хрящевых рыб — и приспособления для прикрепления яйца к субстрату). Яйца анамний содержат умеренное количество желтка, испытывают полное, но неравномерное дробление; формирование зародышевых слоев, полости тела и внутренних органов идет у них сходно с эмбриональным развитием бесчерепных (см. выше). Из яйца вылупляется личинка, ведущая водный образ жизни и более или менее похожая на взрослое животное. Сильно отличается строение личинок бесхвостых амфибий, которые лишь путем сложной перестройки — метаморфоза — приобретают признаки взрослых животных.
Для амниот (первично-наземных животных) характерно усложнение строения яйца (увеличение количества желтка и белка, образование плотных наружных оболочек), внутреннее оплодотворение и способность яйца развиваться только в воздушной среде. Это достигнуто изменением хода эмбрионального развития. На сильно перегруженном желтком яйце делится только анимальный полюс и образуется плавающий на желтке однослойный зародышевый диск. На нем возникает первичная бороздка, через которую часть эктодермальных клеток перемещается под эктодерму, давая начало эндодермальному и мезодермальному слоям. Далее идет образование сомитов и обособление всех внутренних органов. Из краевых участков зародышевого диска формируются специальные зародышевые оболочки, внутренняя из них — амнион — выделяет амниотическую жидкость, в которую оказывается погруженным тело зародыша. Как вырост задней части первичной кишки развивается аллантоис, или зародышевый мочевой пузырь, служащий зародышевым органом дыхания.
Полость тела. Полость тела — целом — выстлана тонкой эпителиальной оболочкой — брюшиной (peritoneum): покрывающая внешние стенки полости тела, называется париетальным листком, а покрывающая внутренние органы — висцеральным листком. На двухслойной брыжейке (mesenterium) как бы подвешены к спинной стороне полости тела внутренние органы, в том числе и пищеварительный тракт. В эмбриогенезе всех позвоночных от передней части полости тела обособляется особая околосердечная полость, в которой лежит сердце; ее оболочка называется околосердечной сумкой (pericardium). У млекопитающих диафрагма делит полость тела на две половины: грудную, в которой лежат легкие и окруженное околосердечной сумкой сердце, и брюшную, где расположены желудок, кишечник, печень, почки, яичники и др. У предков хордовых целомическая полость впервые возникала как опорное образование. У хордовых в связи с образованием миохорда целом потерял опорную функцию, но сохранил рессорное значение, уменьшая опасность повреждения внутренних органов при движении.
Еще по теме Характеристика подтипа:
- ПОДТИП 1. БЕСЧЕРЕПНЫЕ —ACRANIA
- 13.5.7. Подтип Позвоночные Vertebrata
- ПОДТИП 2. ОБОЛОЧНИКИ, ИЛИ ЛИЧИНОЧНОХОРДОВЫЕ,— TUNICATA, SEU UROCHORDATA
- ПОДТИП 3. ПОЗВОНОЧНЫЕ, ИЛИ ЧЕРЕПНЫЕ — VERTEBRATA, SEU С RAN ЮТА
- ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
- ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
- Характеристика экоморф сайгака
- СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ВАКЦИН И РЕКОМЕНДАЦИИ ПО ИХ ПРИМЕНЕНИЮ
- ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
- ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
- ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
- ХАРАКТЕРИСТИКА ТЕПЛА КАК ЭКОЛОГИЧЕСКОГО ФАКТОРА