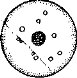
КЛАССИФИКАЦИЯ ЯИЦ ПО КОЛИЧЕСТВУ ЖЕЛТКА И РАСПРЕДЕЛЕНИЮ ЕГО В ЦИТОПЛАЗМЕ
Большинство допросов о причинах дробления-.остается ещепе- ясным из-за недостатка биофизических и биохимических данных. Далека от ясности центральная проблема — почему онтогенез клеток и бластомеров завершается митозом. При такой ситуации нельзя пренебрегать различными гипотезами, которые хотя бы отчасти помогают объяснять процессы дробления. Важнейшей является гипотеза, высказанная О. Гертвигом. Она связывает осо- бенности .дробления с количеством желтка _и разным распределением его в цитоплазме. Вот почему, прежде чем описывать морфологию начальных стадий развития зародышей разных животных, мы должны остановиться на классификации яиц по количеству и распределению желтка.
Алецитальные яйца. Это яйца почти без желтка или с. очень незначительным количеством его. -Fакопы, "например, яйцевые клетки [млекопитающих, плоских червей (рис. 29,Д). Поскольку яйцевые клетки млекопитающих все же содержат небольшое ко' личество относительно равномерно расположенного желтка, то их можно называть и изолецитальными яйцами.
Гомолецитальные, или извлек цитальные, яйца. В этих яйцах желток относительно равномерно распределен в цитоплазме.^дро у таких яиц расположено, в центре (рис. 29,Б). Такое строение Имеют яйца многих моллюсков, иглокожих и ланцетника. Гомо- ‘лецитальные яйца могут иметь и очень большое количество желтка, например яйца гидр или не- мертины Lirieus gesserensis.


 Телолецитальные яйца (рис.
Телолецитальные яйца (рис.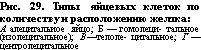 29,Б). Желто к редко неравномерно распределен в цитоплазме Й области аномального полюса мало или почти совсем нет желтка; большая часть его находится в вегетативном полушарии, где желточные зерна и•пластинки лежат более плотно.
29,Б). Желто к редко неравномерно распределен в цитоплазме Й области аномального полюса мало или почти совсем нет желтка; большая часть его находится в вегетативном полушарии, где желточные зерна и•пластинки лежат более плотно.Примером телолецитальных яиц являются яйца амфибий, на- пример лягушек. Ооцит и зрелое яйцо лягушки отчетливо поляр- ,ны: овд~1|мрйГ длинную ось и_ перпендикулярно к ней — более короткую. В области вехетативного‘4юлю^яШ10^ёветлоё..,(сёрое или желтоватое), ацимальная же половина пигментирована. У яиц одних видов лягушек пигмент черный, а у других —- 'бурый или коричневый. Отмечен и градиент расположения РНК вдоль ани-
мально-вегетативноц оси Положение ядра эксцентричное; оно
находится- в анимальиой половинёГвыше экватора. В анимальиой половине значительно .меньше желтта^~чбм“Т^егетат'йвнЖПЕсли мы повернем оплодотворенное яйцо так, чтобьг пигментированный полюс оказался внизу, а светлый наверху, то яйцо незамедлительно возвратится к нормальному положению. Это обусловлено обилием желтка в вегетативном полушарии, имеющем больший удельный вес по сравнению с остальными частями яйца.
Такое же строение имеют яйца птиц (см. рис. 3), у которых, однако, область желтка достигает ,огромных размеров по сравнению с очень небольшим участкдк.'^аС-ЗЩ^^шёвйй' ‘ дйсКд'м), менее загруженным желтком. Здесь расположено ядро. Не случайно собственно яйцо птицы в общежитии называется желтком. Зародышевый диск в первые дни развития зародыша находится наверху. Это имеет, вероятно, большое физиологическое значение, так как «верхняя часть» яйца под наседкой всегда имеет более высокую температуру, чем нижняя (выше на 2—3 и более градусов). Обычная создающаяся под наседкой температура оказывается оптимальной для собственно зародыша, но она является крити
ческой для различных ферментов, имеющихся в вегетативном полушарии.
Телолецитальные яйца с огромным количеством желтка, занимающим большую частgt;'яйца, свойственны и некоторым другим животным, например головоногим моллюскам.
Центролецитальные яйца имеют большое количество равномерно расположенного желтка (поэтому могут быть условно названы также гомолецитальными), однако у поверхности яйца имеется тонкий слой цитоплазмы, почти лишенной желтка (рис. 29,Г). Расположенное ближе к центру яйца ядро также окружено слоем относительно чистой цитоплазмы. Эти яйца характерны для членистоногих.
МОРФОЛОГИЯ ДРОБЛЕНИЯ ЯИЦ
Полное и частичное дробление. Различают, дробление полное или, голобластическое, и частичное — меробластическое.
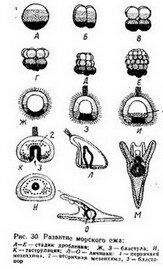 При полном дроблении (рис. 30) вся яйцевая клетка дробится на меньшие клетки. Проникая до самых глубоких частей яйца, борозды дробления распространяются в то же время по всей поверхности яйца. Это свойственно яйцам але- цитального, изолецитальцо- го типа й некоторым яйцевым клеткам телолёциталь- ного типа, например яйцам амфибий (см. рис. 33).
При полном дроблении (рис. 30) вся яйцевая клетка дробится на меньшие клетки. Проникая до самых глубоких частей яйца, борозды дробления распространяются в то же время по всей поверхности яйца. Это свойственно яйцам але- цитального, изолецитальцо- го типа й некоторым яйцевым клеткам телолёциталь- ного типа, например яйцам амфибий (см. рис. 33).При частичном дроблении борозды дробления не проникают глубоко внутрь яйца, большая часть яйца оказывается вне сегментации. Частичное дробление, в свою очередь, у разных животных оказывается очень своеобразным. Следует_раз.-_ личать поверхностное и.ди- скоидальное дробления. Поверхностное дробление (рис. 31), характерное для насекомых и большинства других членистоногих, отличается следующими особенностями, связанными сострук-

реющие на поверхность яйца ядра располагаются на довольно близких расстояниях друг от друга, образуя синцитиальный слой. Затем наступает процесс очень своеобразной" сегментации периферийной, цитоплазмы, происходит постепенное обособление вокруг ядер клеточных территорий^ (рис. 31,В, Б). Борозды дробления появляются, но они не заходят глубоко в яйцо. ЕГрё- зультате периферийная часть яйца превращается в сплошной эпш телиеподобный слой клеток—в бластодерму, внутренние участки которой не ограничены от нераздробившейся центральной массы яйца. На следующем этапе развития зародыша бластодерма мор- фодогически целиком обособляется от желтка и приобретает ис- тинно клеточную структуру. У большинства яиц, которым свойственно такое дробление,, отдельные ядра С ' небольшим участком цитоплазмы-.остаются в скоплениях желтка. Эти клетки — вит ел- лофаги («пожиратели желтка», желточные клетки), перерабаты-, вающие желток, не участвуют непосредственно в построении зародыша, но выполняют важную физиологическую роль в~э11брй- ональном развитии.

При развитии центролецитальных яиц полость дробления не возникает, и только на более поздних стадйяЗГ'фазвития бласто- Дерма. отделяется ют лежащетдод ней желтка.
У многих яиц, которым свойственно также частичное дробление, оно совершается не по всей поверхности, а лишь в районе
анимального полюса, где расположено ядро. Такое дробление пашваетсяГШЬшШ^ь'ньш. Оно особенно распространено""^"по- зводочных и характерно для акул, скатов, костистых рыб;" пресмыкающихся, иптиц (рис. 32).
Равномерное и неравномерное дробление. Образующиеся в результате дробления бластомеды могут казаться почти одинаковыми по размерам. Такое дробление называется равномерным. Оно свойственно,..как. правил о, гомодецитальным яйцам.
Более распространено неравномерное дробление. Типичным
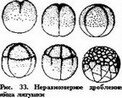

примером является дробление яйца лягушки, начиная с третьего дробления (рисГЗЗу.
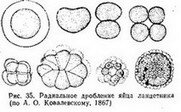
Строго говоря, настоящего равномерного дробления ие существует. Уже первые два Эластомера при любом характере дробления отличаются друг от друга и по размерам и, особенно, по структуре й биохимическим свойствам (см. гл. XI). В отношении некоторых животных, например ланцетника, гово
рят, что дробление их яиц почти равномерное. По внешнему виду бластомеры кажутся одинаковыми, на самом же деле в области вегетативного полюса бластомеры большего размера и отличаются по форме. Яйцо одного и того же вида животного на разных этапах развития может дробиться равномерно или неравномерно. Например, первые два дробления яйца лягушки (рис. 33), равномерные, а последующие — резко неравномерные.
 Синхронное и несинхронное дробление. Последовательные дробления бластомеров могут .щк^гущать..одновременно,, например у яиц морских' ежей до 32-бластомерной стадии или у амфибий на ранних этапах дробления. Такое синхронное дробление отличается от несинхронного дробления, которое наблюдается у низших многоклеточных :Живбтных,.у некоторых червей и млекопитающих.
Синхронное и несинхронное дробление. Последовательные дробления бластомеров могут .щк^гущать..одновременно,, например у яиц морских' ежей до 32-бластомерной стадии или у амфибий на ранних этапах дробления. Такое синхронное дробление отличается от несинхронного дробления, которое наблюдается у низших многоклеточных :Живбтных,.у некоторых червей и млекопитающих.Классификация дробления, основанная на взаимном пространственном расположении бластомеров. В этой классификации выделяют несколько типов.
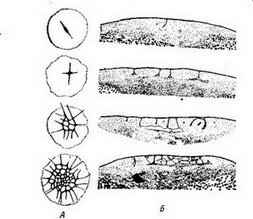
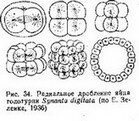
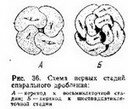
Радиальное дробление (рис. 34 и 35) в его наиболее типичном виде характерйзубтСя~еЛёдующими особенностями. Первое дробление меридиональное^ оно проходит в меридиональной плоскости яйца. Второе дробление также меридиональное, оно проходит через главную ось яйца, нр под прямым углом к плоскости первого дробления. Плоскость 3-го дробления, лежит под прямым углом
дробление происходит в плоскости, пар аллельной" экв атсфу, поче

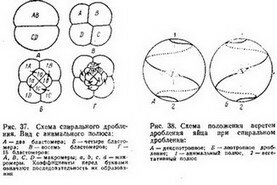
Для аннелид, большинства моллюсков, немертин и некоторых планарий (Polyclada) характерно спйуальндё дтй'лениё (рис. 36 и37У7"При этом типе дробления бластомеры, входящие в состав горизонтальных рядов (образовавшиеся в результате экваториаль- Делений), не располагаются один над другим: бластомёры
верхнего рида..как бы вклиниваются между бластомерами нижнего ряда.
мьга углом к преды-. дущйм и, значит, опять-таки под углом в 45° к экватору. Если мысленно продолжить веретено дробления в обе стороны по поверхности яйца, то получится спиральная линия (рис. 38). Такая ситуация создается благодаря тому, что в анафазе каждого деления дробления два дочерних бластомера отклоняются от оси веретена в противоположные стороны (В. Н. Мещеряков).
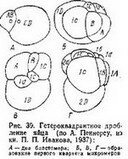
 После первых двух дроблений образуются четыре бластомера, которые называются основным квартетом и обозначаются А, В, С в. D. Бластомеры основного квартета могут быть примерно одинакового размера (рис. 37,Б), тогда образующиеся при последующих дроблениях более мелкие клетки в анимальной области будут также одинаковой величины (рис. 37,В). Такое дробление принято называть гомоквадрантным. Если клетки основного квартета не одинаковы по величине, то и их производные также различаются. Обычно самые крупные размеры имеет бластомер D и соответственно микромеры, которые от него отделятся. Неравномерное спиральное дробление называют гетероквадрантным (рис. 39).
После первых двух дроблений образуются четыре бластомера, которые называются основным квартетом и обозначаются А, В, С в. D. Бластомеры основного квартета могут быть примерно одинакового размера (рис. 37,Б), тогда образующиеся при последующих дроблениях более мелкие клетки в анимальной области будут также одинаковой величины (рис. 37,В). Такое дробление принято называть гомоквадрантным. Если клетки основного квартета не одинаковы по величине, то и их производные также различаются. Обычно самые крупные размеры имеет бластомер D и соответственно микромеры, которые от него отделятся. Неравномерное спиральное дробление называют гетероквадрантным (рис. 39).В зависимости от направления смещения мелких бластомеров (микромеров) по отношению к крупным бластомерам (макромерам) спиральное дробление подразделяется на два типа. При дексиотропном дроблении смещение микромеров осуществляется по часовой стрелке (если смотреть на дробящееся яйцо с анималь- ного полюса). Такой тип дробления изображен на рис. 36 и 37. При леотропном дроблении микромеры смещаются против часовой стрелки. Первые четыре микромера называют первым квартетом микромеров. Их обозначают малыми буквами латинского алфавита с коэффициентом 1 (la, lb, lc, Id). Основной квартет после выделения первого квартета микромеров называют первым квартетом макромеров; обозначаются они соответственно 1А, 1В, 1C и 1D. При образовании квартетов микромеров происходит передо-
вание дексиотропных и леотропных дроблений. Для возникающих после четвертого дробления 16 бластомеров приняты такие индексы: бластомер первого квартета микромеров 1а дробится на бластомеры 1а1 и 1а2, а бластомер основного квартета 1А дробится на бластомер 2а и 2А. Бластомеры 2а, 2Ь, 2с,и 2d составляют второй квартет микромеров, а бластомеры 2А, 2В, 2С и 2D образуют квартет макромеров. Возникшие в результате очередных дексиотропных и леотропных дроблений 32 и 64 бластомера обозначаются по тому же принципу.
В ходе спирального дробления чаще всего выделяется четыре квартета микромеров, которые соответственно занимают все более удаленное положение от ани- мального полюса. Поскольку одновременно с делением макромеров делятся и первые квартеты микромеров, то к концу дробления на анимальном полюсе образуется значительное количество клеток— производных первого квартета
микромеров. Поскольку общее число клеток, образующихся при спиральном дроблении, невелико, то удается проследить «судьбу» каждого бластомера. При использовании буквенно-цифровой номенклатуры все бластомеры получают определенный индекс. Оказалось, что все бластомеры, обозначаемые малыми буквами и имеющие коэффициент 1, т. е. все клетки, образовавшиеся из первого квартета микромеров, пойдут на построение предротового отдела личинки. Из клеток второго квартета возникает венчик ресниц — прототрох; третий квартет микромеров пойдет на образование мезодермальных и отчасти эктодермальных производных личинки и, наконец, четвертый квартет микромеров и четвертый квартет макромеров образуют кишечник. Особую роль при спиральном дроблении играют бластомеры 2d и 4d. Первый называется соматобластом, и из него образуется эктодерма заднего отдела личинки, а в дальнейшем покровы взрослого червя, 4d называется мезобластом. Этот бластомер делится на две клетки телобласты — родоначальницы мезодермальных телобластических полосок, из которых образуются целомы личинки, а в ряде случаев и целомическая мезодерма взрослого организма.
По мнению О. М. Ивановой-Казас, спиральное дробление развивалось постепенно. Элементы этого типа дробления встречаются у низших многоклеточных. Так же постепенно происходило и его «вырождение», т. е. замена другими формами дробления.
Дробление яиц нематод, коловраток, асцидий и некоторых других животных имеет ту особенность, что уже на ранних этапах дробления проявляется билатеральная симметрия в расположении бластомеров (см. рис. 8). Такое дробление называется билатеральным или двустороннесимметричным. Вне зависимости от своеобразия дробления в деталях (у разных животных) при билатеральном типе дробления каждый бластомер правой половины эмбриона соответствует подобному бластомеру левой половины.
Следует упомянуть об очень своеобразном двусимметричном типе дробления, свойственном ктенофорам, у которых уже на стадии четырех бластомеров определяются обе плоскости симметрии будущего организма — продольная и поперечная.
Строго закономерный характер дробления яиц каждого вида животных обусловлен сложившимся в ходе эволюции особым морфофизиологическим механизмом. Кажущимся исключением является анархическое дробление, которое иначе называют «беспорядочным» и «хаотическим», например дробление яиц некоторых метагенетических медуз. Анархическое дробление гидромедузы Oceania .armata (рис. 40) описал впервые И. И. Мечников в 1886 г. Как и у других гидромедуз, никакой «анархии» в начале дробления нет: первая борозда дробления меридиональная, врезающаяся; появляется она на анимальном полюсе; вторая борозда тоже меридиональная, но появляется на внутренней стороне бластомеров и распространяется центробежно (рис. 40,Б). Однако
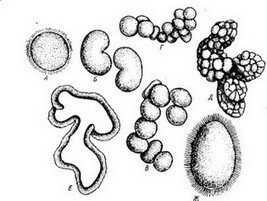
Рис. 40. Развитие гидромедузы Oceania armaia.
А—Г — дробление; Д—Е — бластула; Ж — личинка (по И. И. Мечникову, 1886)
порядок взаимного расположения бластомеров после третьего дробления может варьировать. Чем дальше идет дробление, тем •более проявляется «беспорядочный характер» (рис. 40,В, Г).
Такие своеобразные явления дали повод П. П. Иванову (1945) утверждать, что яйца некоторых кишечнополостных не имеют никакой детерминации и предварительной структуры. Это утверждение трудно аргументировать, так как анархический тип дробления кажется хаотичным лишь при условии, если закономерным считают только такое пространственное расположение бластоме- .ров, которое укладывается в те или иные принятые при описании дробления геометрические схемы, в понятие о симметрии. Совершенно очевидно, однако, что при кажущемся совершенно «беспорядочным» дроблении яйца гидромедуз развиваются в животные именно данного, а не другого вида. Не выяснены еще полностью закономерности такого развития.
Кажущаяся хаотичность и простота анархического дробления позволили Л. Н. Жинкину в 1951 г. высказать предположение о том, что оно является примитивным, не установившимся и что ют него произошли спиральный и радиальный типы дробления.
Черты спирального дробления у кишечнополостных отмечал ещ И. И. Мечников в 1886 г., а впоследствии С. Берг (1941) и К. Ни хольм (1949). По гипотезе Г. А. Шмидта (1951), радиально дробление возникло из спирального.
У актинии Sagartia troglodytes дробление может варьировать Оно может быть равномерным или неравномерным, бластомерь могут располагаться как при радиальном, так и при спирально» дроблении.
Вопросы эволюции дробления пока еще слабо разработаны Вероятно, будут иметь значение исследования эволюции дробле ния в пределах отдельных типов. При анализе анархическогс дробления следует принять во внимание своеобразную способность к полиэмбрионии и соматическому эмбриогенезу (см гл. XVI) зародышей многих кишечнополостных. Фрагменты зародыша гидромедузы Oceania armata способны развиваться в нормальные личинки уменьшенного размера. Подобные явления отмечены и у гидр. М. Н. Грузова, изучая развитие гидр, в 1956 г. обнаружила, что зародыши, пройдя нормальное, закономерное развитие вплоть до гаструляции, при формировании эмбриотеки (особая оболочка вокруг зародыша) и все зимнее время оказывается менее дифференцированным, анархизированным и напоминает структуры геммул и сомитов (см. гл. XVI), хотя до формирования эмбриотеки развитие происходит строго закономерно. У гидр дробление почти полное и равномерное. Оно заканчивается целобластулой (см. далее), и закономерно совершается гаструля- ция. И вот при таком закономерном развитии зародыш теряет свою нормальную интеграцию и в некоторых случаях вместо одного нормального зародыша развиваются два. Предполагают, что в результате такой полиэмбрионии начинается развитие по типу соматического эмбриогенеза, еще недостаточно изученное эмбриологами (см. гл. XVI).
Интересно в этой связи отметить, что у Oceania полиэмбрионии не обязательно имеет место, не все бластулы делятся. Форма некоторых эмбрионов становится постепенно все более правильной, зародыш удлиняется и, наконец, принимает вид языкообразной планулы. Это означает, что нет анархии в дроблении, но имеется стадия дезинтеграции зародыша, которая может привести к полиэмбрионии.
Как видим, ничто не свидетельствует о хаосе анархического дробдения, и этот термин нельзя считать удачным. Многие водные низшие многоклеточные организмы обладают низким уровнем интеграции; яйца их мало отличаются от соматических клеток (они не имеют обычных яйцевых оболочек, у них относительно нормальное, как и соматических клеток, соотношение ядра и цитоплазмы, нормальный метаболизм). У этих малоинтегрированных организмов и их зародышей выработался особый характер дробления как адаптация к меняющимся внешним условиям, выработалась возможность перехода к соматическому эмбриогенезу. Более подходит к такому типу дробления название регуляционный тип дробления.
Источник: Токин Б. П.. Общая эмбриология: Учеб, для биол. спец, ун-тов.—4-е изд., перераб. и доп. 1987
Еще по теме КЛАССИФИКАЦИЯ ЯИЦ ПО КОЛИЧЕСТВУ ЖЕЛТКА И РАСПРЕДЕЛЕНИЮ ЕГО В ЦИТОПЛАЗМЕ:
- Количественный метод копрооволарвоскопии и подсчет количества яиц и личинок гельминтов в г фекалий
- СОЗРЕВАНИЕ СПЕРМИЕВ (СПЕРМИОГЕНЕЗ) И ЯИЦ (ОВОГЕНЕЗ)
- СОЗРЕВАНИЕ СПЕРМИЕВ (СПЕРМИОГЕНЕЗ) И ЯИЦ (ОВОГЕНЕЗ)
- 7.3. МОРФОФИЗИОЛОГИЧЕСКИЕ И ЭВОЛЮЦИОННЫЕ ОСОБЕННОСТИ ЯИЦ ХОРДОВЫХ
- Значение количества пищи
- Разработка метода, определения остаточных количеств препаратов.
- ОПРЕДЕЛЕНИЕ ОБЩЕГО КОЛИЧЕСТВА ЛЕТУЧИХ ЖИРНЫХ КИСЛОТ (ЛЖК)
- ГЛАВА ДЕСЯТАЯ Подтверждение опытами первой предпосылки о количестве переходящей из вен в артерии крови и опровержение возражений
- Распределение активности во времени суток
- Распределение и депонирование токсикантов в организме
- Общие принципы распределения токсикантов в организме.
- РАСПРЕДЕЛЕНИЕ МИКРООРГАНИЗМОВПО ПОЧВЕННОМУ ПРОФИЛЮИ ИХ ПЕРЕМЕЩЕНИЕ
- РАСПРЕДЕЛЕНИЕ ЭЛЕМЕНТОВ В РАЗЛИЧНЫХ КОМПОНЕНТАХ ЭКОСИСТЕМ СУШИ