Случайное размещение
Такое размещение возникает в том случае, если среда более или менее равномерна и особи не испытывают ни притяжения друг к другу, ни отталкивания. Именно так распределены в муке при сравнительно небольшой плотности популяции личинки большого мучного хрущака Tenebrio molitor L., а также личинки жуков-стафилинид на поверхности почвы.
Остановимся далее на моделировании случайного размещения. Биномиальное распределение, рассмотренное в предыдущем разделе, предусматривает достаточно высокую плотность популяции, когда число незанятых территорий относительно мало. Если же изучаемый объект редок (р ® 0) и распределен по потенциальным территориям случайным образом, а само число этих территорий велико (n ), то биномиальное распределение переходит в распределение редких событий (распределение Пуассона). При этом вероятность обнаружения в пробе того или иного количества насекомых будет зависеть только от размера пробы, но не от того, где именно эта проба взята.
Согласно формуле Пуассона, ожидаемое число проб nx с количеством особей x равно:
nx = ^Nem x!
где N - общее количество всех взятых проб, m - среднее число особей на одну пробу, е - основание натуральных логарифмов (е @ 2,71828) , х!- икс-факториал - произведение ряда натуральных чисел, например 3! = 1-2-3.
В случае распределения Пуассона дисперсия и арифметическая средняя примерно равны, т.е. 82 / m @ 1
Если же вероятность нахождения объекта достаточно высока, то упомянутое выше биномиальное распределение превращается в широко известное из статистики нормальное распределение (распределение Гаусса). В переходных случаях максимум кривой распределения обычно смещен в сторону меньших значений признака (влево), так как относительно велико число проб с небольшим количеством особей (рис.21). При малых же пробах максимум кривой распределения смещается на 0 (т.е. большинство проб - пустые) и от кривой остается только нисходящая ветвь.
Агрегированное размещение
Размещение этого типа наиболее обычно в природе. Насекомые скапливаются на участках, где для них особенно благоприятен микроклимат, имеются их кормовые растения или жертвы. Кроме того, многие насекомые привлекают друг друга благодаря наличию так называемых агрегационных феромонов (М.Барбье, 1978). Степень агрегированности может зависеть от погоды, сезона и времени суток. Насекомые в малоактивном состоянии чаще образуют скопления в благоприятных для переживания местах. Последнее в первую очередь относится к хищным насекомым.
![Рис.21. Пуассоновское распределение вероятности (по А. А. Шарову, 1984)]()
Рис.21. Пуассоновское распределение вероятности (по А. А. Шарову, 1984)
Агрегированное пространственное размещение особей может быть различных типов, а именно: группы могут быть сходными или различными по численности, размещение особей в пределах группы является или случайным, или равномерным. Кроме того, в ряде случаев группы могут объединяться в скопления групп. Соответственно, эти размещения описываются различными теоретическими кривыми. В любом случае, однако, здесь коэффициент дисперсии всегда больше единицы.
Один из конкретных примеров агрегированного размещения дают личинки комаров- долгоножек, обитающие в почве. Они скапливаются группами примерно по 30 особей. На каждую особь приходится по 1-2 см2, а на 1 м2 можно найти около 10 таких скоплений. Отдельные особи могут оказываться и в пространстве между группами.
Как уже отмечалось выше, уровень агрегированности популяции насекомых можно определить двумя путями, либо с помощью индексов агрегированности, показывающих степень неравномерности размещения, либо проверкой соответствия данного размещения определенному теоретическому распределению, что, конечно, является более точным описанием.
Однако применение того или другого способа будет корректным только после предварительного анализа размещения.
Дело в том, что полученная нами картина распределения может существенно зависеть от размера пробы, в частности, от того, меньше или больше она, чем средний размер скоплений.
Для проверки целесообразно взять серии проб разного размера, например, при исследовании почвенной фауны 40 проб по 0,25 м2, 20 проб по 0,5 м2 и 10 проб по 1 м2 (А.В.Смуров, 1975). Для каждой из серий необходимо определить среднеарифметическую величину сбора данных насекомых на одну пробу (m) и дисперсию (8 2 ).
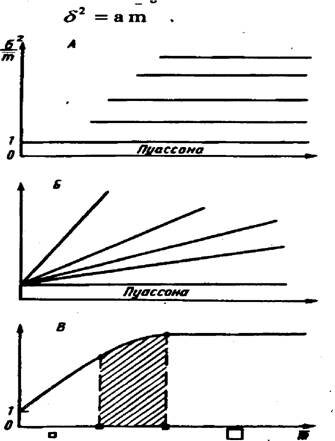
По виду зависимости дисперсии от средней можно выбрать индекс агрегированности и теоретическое распределение для описания размещения насекомых. В одном случае коэффициент дисперсии (дисперсия, деленная на среднеарифметическую) не будет зависеть от средней, т.е. будет почти одной и той же величиной, независимо от размера пробы, в другом же - будет сначала линейно возрастать с увеличением средней (размера пробы), отходя от единицы на оси ординат (рис.22). При достижении пробой величины скопления насекомых на графике имеет место точка перегиба кривой и далее с увеличением пробы коэффициент дисперсии почти не меняется.
В первом случае в качестве индекса агрегированности можно выбрать сам коэффициент дисперсии. Теоретические распределения, соответствующие данному случаю - распределения Неймана (J.Neyman, 1939) и Томас (M.Thomas, 1949). Эти распределения имеют место, когда размер пробы значительно больше скопления насекомых. В общем такие случаи редки в энтомологической практике и мы не будем здесь далее на них останавливаться.
Гораздо чаще коэффициент дисперсии возрастает с увеличением пробы, т.е. скопления велики по сравнению с площадью, охватываемой пробой. В этом случае необходимо использовать иные, чем коэффициент дисперсии, индексы агрегированности. Перечислим некоторые из них.
Л.Р.Тейлор (L.R.Taylor, 1961) на основании изучения размещения на местности многих видов насекомых предложил следующую эмпирическую формулу:
82 = ашъ
![Рис.22.<div class=]()
Зависимость коэффициента дисперсии от средней для ряда реальных размещений, описываемых различными теоретическими распределениями: А - Неймана и Томас; Б - отрицательным биномиальным и трехпараметрическим; В - гипотетическим реальным при непрерывном увеличении размера пробы и соответственном увеличении средней." />
Рис.22. Зависимость коэффициента дисперсии от средней для ряда реальных размещений, описываемых различными теоретическими распределениями: А - Неймана и Томас; Б - отрицательным биномиальным и
трехпараметрическим; В - гипотетическим реальным при непрерывном увеличении размера пробы и соответственном увеличении средней.
Квадратами разного размера под осью абсцисс показаны относительные размеры проб. При малом размере проб здесь возможно использовать отрицательное биномиальное и трехпараметрическое распределения, при большом - распределения Неймана и Томас. При средних размерах пробы теоретическое распределение не может быть подобрано (заштрихованная область) (по А. В. Смурову, 1975)
40
где а - величина, зависимая от размера пробы, а b - индекс уровня агрегированности. Чем больше b, тем выше этот уровень. Коэффициенты а и b можно вычислить, логарифмируя приведенное выше уравнение:
log 82 =log a + b log m .
Так как это уравнение с двумя неизвестными, необходимо провести две серии учетов с разными размерами проб.
Другая тоже эмпирическая формула была предложена японскими исследователями (S.Ivao, E.Kuno, 1968):
82 = (b - 1)m2 + (a + 1)m
В этой формуле коэффициент a определяет, что является основным параметром анализируемого размещения: отдельные особи или их скопления, а коэффициент в показывает агрегированность этих компонентов в пространстве.
Чаще, по крайней мере в отечественной литературе, используют тоже эмпирический индекс агрегированности М. Мориситы (M.Morisita, 1959):
Sqx,(x,-1) q N(N-1) '
где I - индекс агрегированности, q - общее число проб данного размера, х,- реальное число особей в каждой пробе, N - общее (суммарное) число особей во всех q пробах.
Этот показатель при агрегированном размещении больше единицы.
Агрегированное размещение во многих случаях хорошо описывается, так называемым отрицательным биномиальным распределением. Поэтому в качестве индекса целесообразно использовать константу К этого распределения. Ее можно подсчитать по следующей формуле:
82 - 1т
— = m — +1
m K
Приближение К к нулю свидетельствует о большой агрегированности популяции. Наоборот, чем больше К, тем распределение сборов по пробам ближе к случайному. Так, по данным В.Е. Уотерса (V.E.Waters, 1959), при низкой плотности популяции Choristoneura fumiferana константа К изменяется в пределах 1-5 (агрегированное размещение). При более высокой плотности популяции их размещение становится все более случайным (значение К в пределах 6 - 50).
Иногда используют также величину С, обратную К:
С = 1 = 82 1 = 82 - m
2 2
K m m m
Величина С может быть определена как тангенс угла наклона графика зависимости коэффициента дисперсии от средней (от размера пробы).
Индекс Мориситы и константы отрицательного биномиального распределения связаны друг с другом следующим образом:
I = C +1 = — +1 K
А.В.Смуров (1975) предложил в качестве индекса агрегированности KA следующую формулу:
KA=1-m
m
где m- средняя плотность популяции в пределах всей занимаемой ею территории, включая незаселенные пространства, m - средняя плотность в местах скоплений. При полностью случайном распределении этот индекс равен нулю, а при максимальной агрегированности он будет стремиться к единице. Индекс А.В.Смурова имеет наглядный биологический смысл, так как показывает, какая часть общей площади не занята скоплениями. Связь между этим индексом и предыдущими можно выразить следующим образом:
K. =1-1= 1
I K +1
Мы уже упоминали выше, что в случае агрегированного размещения особей коэффициент дисперсии сначала увеличивается при увеличении размера пробы (соответственно и увеличении среднего количества особей в пробе). Эти изменения связаны с соотношением размера пробы и средней площади, занимаемой скоплением особей.
В принципе здесь мы всегда имеем дело, как минимум, с "двойным" размещением: размещением скоплений на местности и размещением самих особей внутри скоплений. Возможно и образование групп скоплений.
Известны два теоретических распределения, соответствующих данной ситуации. Это отрицательное биномиальное распределение (CJBliss, R.A.Fischer, 1953; рис.23) и трехпараметрическое распределение (А. В. Смуров, 1975). На практике чаще используют сравнение эмпирических данных с расчетом по формуле отрицательного биномиального распределения (эта формула является лишь хорошей апроксимацией, более подробно это изложено в книге А.В.Смурова и Л.В.Полищука, 1989) Согласно формуле отрицательного биномиального распределения вероятность обнаружения в пробе x особей
P = 4K+x) /_m_Y. (_A
где Г(К+х) и Г(К) - гамма-функции, т.е. факториалы дробных чисел, К - константа отрицательного биномиального распределения, а m - среднее количество особей в пробах. Гамма-функция может быть вычислена по следующей приближенной формуле:
Г^) = 72p(x -1) • (x - 1)(x-1) • e-x+1.
Конкретный пример расчета отрицательного биномиального распределения дан
А.А.Захаровым и др. (1987) (На стр.204 цитируемой книги допущена опечатка - вместо м = (xp)/(I - р) следует читать м = (xp)/(і - р)). Как уже отмечалось выше, А.В.Смуровым было предложено так называемое трехпараметрическое распределение, основанное на следующих параметрах: общей средней плотности популяции, плотности в пределах скоплений и плотности вне скоплений. Согласно этому распределению, как в случае отрицательного биномиального распределения, коэффициент дисперсии также меняется в зависимости от величины пробы.
![Рис.23 Отрицательное биноминальное распределение вероятности (по А. А.Шарову, 1984)]()
Рис.23 Отрицательное биноминальное распределение вероятности (по А. А.Шарову, 1984)
Все эти теоретические распределения могут переходить друг в друга при существенном изменении их параметров. Поэтому, если нет уверенности в справедливости какой-либо определенной модели распределения, рекомендуется использовать так называемое трехпараметрическое распределение Пуассона с нулями (А.В. Голубев, Г.Э. Инсаров, В.В. Страхов, 1980), являющееся наиболее универсальным. 3.