Продукция биомассы
Когда растения растут, увеличивая при этом за время роста количество биомассы на единицу площади, говорят о продукции биомассы, или (выражая интенсивность за единицу времени) о продуктивности.
Поскольку продукция биомассы растений стоит в основании пищевой цепи, ее называют первичной продукцией. Различают брут- то-первичную продукцию ВРР, т. е. количество биомассы, которое в совокупности синтезируется на единицу площади, и нет- то-первичную продукцию NPP, которая результируется после вычета текущих потерь на дыхание R в экосистеме:NPP - ВРР - 5Ж
Эти продукционные термины, известные из хозяйственной жизни и употребляемые в биологии вообще в большой степени, имеют теоретическую сущность и на практике едва ли употребимы, поскольку величины потерь, в особенности тех, которые происходят в подземном слое и через дыхание, как правило, неизвестны. Оценки NPP, таким образом, могут грешить большими ошибками.
Как МОЖНО принять из приблизительных контрольных цифр, примерно половина усвоенного растением углерода еще при его жизни отдается как С02, выделяемое при дыхании. Другая часть, как правило, возвращается в атмосферу при микробном разложении. В глобальном сопоставлении данных для лесов Дж. Райх и К. На- дельхоффер продемонстрировали, что ежегодная продукция наземного опада (70 — 500 г/м 2 - а-1) соответствует высвобождению С02 из почвы («почвенному дыханию»), из чего можно заключить, что надземная и подземная продукции определяются одними и теми же факторами.
NPP— величина, употребляемая чаще всего, еще никогда не была определена даже предположительно по вышеприведенной формуле. Для этого следовало бы знать за весь период наблюдения интегральное количество связанного углерода и все потери на дыхание. Этого избегают и обыкновенно рассматривают изменение запасов биомассы (АВ) между двумя временными точками (г • м 2 а-1).
Проблема состоит в том, что в течение интервала наблюдений очень много продуцируемой биомассы снова теряется. Отмершие части побегов (VD) еще можно непрерывно собирать и добавлять в заключительных расчетах (естественно без учета потерь летучих веществ типа изопрена — Ех0); труднее реконструировать ту биомассу, которую «консумировали» фитофаги и патогенные организмы (К„), для подземных же органов совершенно невозможно определить потери и «консум» биомассы (V„, К„). В злаковых сообществах больше двух третей биомассы приходится на подземные органы, и большая часть тонких корней недолговечна. Есть оценки, согласно которым еще 5 — 10% ассимиля- тов попадет в микоризообразующие грибы и там на короткое время задерживаются, что тоже не определимо. Количество корневых выделений в ризосферу (сахара, аминокислоты) в значительной степени неизвестно, как и в большинстве случаев потери растворимых органических соединений (dissolved organic matter — DOM), уходящих в просачивающуюся воду (Ех11). Если NPP, определенная на основе изменений запасов биомассы АВ, может оказаться неправильной на 100%, то уравнениеNPP = АВ + Vc + V„ + К,, + Ku + EXo + Ехи
(V — отмершая, потерянная биомасса; К — кон- сумированная биомасса; Ех — экспорт биомассы как надземный, так и подземный) практически нерешаемо. Только в том случае, если все эти величины потерь во всех экосистемах равновелики, данные по NPP, полученные на основании урожая биомассы (АВ), были бы сравнимы, но это совершенно невероятно. Дополнительная проблема— распределение биомассы в течение периода наблюдений. Поскольку новые надземные структуры, строящиеся из резервов в лежащих глубоко в почве органов, в действительности в течение периода наблюдений не были продуцированы (англ. stored growth), биомасса лишь (с метаболическими затратами) переотлагалась снизу вверх.
Из-за неизвестных величин потерь более корректно вместо NPP говорить о нет- то-приросте фитомассы или, как принято в сельском хозяйстве, об урожае (от англ.
harvested yield). Из-за этой неточности значения NPP цитируются как NPP. Почти 46 — 50% сухого вещества приходится на углеродные единицы. Средняя энергетическая емкость (теплотворная способность) биомассы составляет для наземных растений 18,1 кДж, для океанического планктона — 19,3 — 20,6 кДж.Если прирост биомассы в крупных био- мах Земли оценивают как практикующееся приближение к настоящей «продуктивности», результат зависит от выбранного масштаба времени. Если включать непродуктивное время года в областях, близких к полюсу (зимний покой), т.е. оценивать прирост биомассы за год независимо от фактического времени продуцирования, то продуктивность по направлению к полюсу будет падать. Если сравнивать только периоды активного роста, то продуктивность везде на Земле примерно одинаковая пока достаточно воды. Этот поразительный, часто не замечаемый результат показывает, как исключительно физиологические приспособления повсеместно уравновешивают климатические различия. Широтные
Сравниваются показатели за год (справа жирным шрифтом), включая периоды ростового покоя во внетропических областях, или в пересчете на средний месяц вегетационного периода (прямоугольники слева), что свидетельствует об очень разных результатах (данные только для гумидных областей). Это наглядно показывает, что глобальная разница в годовой аккумуляции биомассы едва ли находится под влиянием температурных условий во время сезона роста. Средние данные продукции для лесов и для злаковых сообществ не отличаются (данные в скобках показывают картину большой региональной и локальной вариабельности)
различия в продуктивности (не говоря о региональных и локальных различиях в условиях роста) почти исключительно являются результатом различной длительности сезона роста; влияние климата в течение этого сезона очень невелико. Если вместо года взять среднемесячные значения периода вегетации, то высокогорные растения умеренной зоны (Альпы) продуцируют биомассы не меньше, чем в среднем влажный тропический лес, которому в этом не уступают и центрально-европейские буковые леса (рис. 13.39). При особо благоприятных условиях роста, как и в условиях интенсивного сельского хозяйства, аккумуляции биомассы в год в экстремальных случаях достигают свыше 5 кг • м А
Трудность при экстраполяции запасов и прироста биомассы состоит в том, что очень трудоемкие анализы проводятся только в немногих пунктах Земли. Понятно, что для таких работ часто выискиваются «красивые» и зрелые насаждения. Поэтому глобальная биомасса, очевидно, значительно ниже ее оценки периода Международной биологической программы (1968—1974). Глобальный запас земной биомассы был определен тогда в 840 Гт углерода. Новые оценки,
Таблица 13.4. Биомассы центрально- европейского смешанного дубовограбового леса
|
Организмы |
Масса сухого вещества, т- га"1 |
|
Зеленые растения |
275 |
|
Листья древесных растений |
4 |
|
Ветви |
30 |
|
Стволы |
240 |
|
Травы |
1 |
|
Животные (наземные) |
gt; 0,0004 (3—5 кг-га-1) |
|
Птицы |
0,0007 |
|
Крупные млекопитающие |
0,0006 |
|
Мелкие млекопитающие |
0,0025 |
|
Насекомые |
? |
|
Почвенные организмы |
ок. 1 |
|
Дождевые черви |
0,5 |
|
Другие почвенные животные |
0,3 |
|
Почвенная флора |
0,3 |
проводившиеся на площадях с «не идеальной» растительностью и оголенными поверхностями, лежат в пределах 560 — 600 Гт углерода (если не обращать внимания на быстрое исчезновение лесов в тропиках, оценивающееся в 1—2 Гт углерода в год, чему противостоит нетто-прирост в умеренной зоне; см. 13.7.6).
Первичные продуценты своей живой и отмершей массой образуют основу для дальнейшей переработки вещества консу- ментами и редуцентами, т.е. для вторичной продукции. Согласно процессам в пищевых цепях растительная масса в сотни
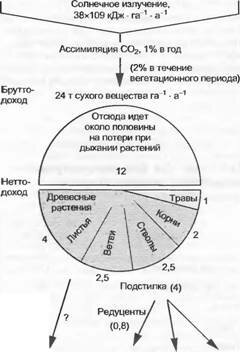
Животные,
поедающие
растения
(lt; 0,02)
Мине-
0,3 0,3 рализи- Гумус
, ї 1 руется 1 0 2 1
Насекомые, Почвенная j Дождевые черви беспозвоночные флора прочие почвенные и др. V / животные
ч х
Хищные животные?
(lt; 0,002)
Рис. 13.40. Годовое излучение Солнца, а также первичная и вторичная продукция в центрально-европейском смешанном дубово-грабовом лесу (ср. табл. 13.4) (по данным P.Duvigne- aud и H.EIIenberg) раз больше, чем масса вторичных продуцентов. Среди консументов наибольшая масса свойственна фитофагам, в то время как хищники первого и второго порядка и соответственно паразиты первого и второго порядка с постепенно уменьшающейся зоомассой составляют вершины пшцевых пирамид с различными трофическими ступенями. Табл. 13.4 и рис. 13.40 иллюстрируют в качестве примера распределение биомасс и величин продукции центрально-европейского дубово-грабового смешанного леса. При этом видно, что пищевая пирамида соответствует пирамиде продукции, поскольку первичная нетто-про- дукция достигает здесь, как и в других экосистемах, на порядок более высокие (в десятки и сотни раз) значения, чем значения вторичной продукции.
Рис. 13.40 показывает также, что консументы принимают лишь незначительное участие во вторичной продукции, потому что из первичной продукции растений в дубово-грабовом лесу непосредственно потребляется фитофагами только около 2 % (в других наземных биоценозах едва ли более 15%, в среднем около 7%). При этом ежегодно почти 25% продукции аккумулирует ся как мертвая органическая субстанция в твердом состоянии (детрит: листовой опад, гумус и др.) или в почвенных растворах (например, гумолигниновые кислоты). Такому количественно важному компартименту этой экосистемы соответствуют большая значимость и продуктивность сапротрофных и минерализующих редуцентов (см. 9.1.1, бокс 11.4); их участие во вторичной продукции составляет 95 %.
Суша
Без lt;50 данных
га о
50- 250- 500- 250 500 1000
Океаны
1000- 1500- 2000-gt;2500 1500 2000 2500
lt;80
80-
120
120- 200- gt;400 200 400
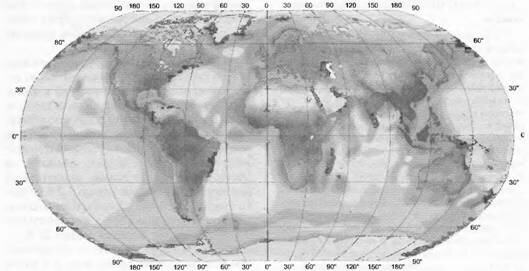
Рис. 13.41. Первичная нетто-продукция биосферы (no Н. Lieth et al., по таким источникам, как РИО- МОДЕЛЬ, а также сайт http:/www.usf.uni-osnabruek.de/hlieth).
Данные приводятся в граммах сухого вещества на 1 м2 в год для суши и океанов
Несмотря на ненадежность выше приведенной базы данных, оценки «NPP» в глобальном сравнении очень иллюстративны (рис. 13.41). Хотя океаны заключают в себе только 0,2 % глобального запаса биомассы, они по своей площади, составляющей 70 % всей поверхности, имеют почти такие же высокие значения «NPP», как и вся суша. Продуктивность тем не менее сосредоточена в богатых питательными веществами областях и регионах, т. е. в прибрежных, с поднимающимися со дна холодными глубинными водами (upwelling regions). Удаленные от побережья тропические и субтропические регионы мирового океана — продуктивные пустыни (белые поверхности на карте). Примечательно, что максимальная естественная продуктивность в море (специфические прибрежные зоны) и на суше может быть очень сходной, а именно 2 000—3 ООО г - м-2- а-1 (точечные пиковые значения «NPP» — до 6000 г м-2 в переходных зонах от суши к морю, почти как в тропических и субтропических болотах). На суше среднегодовая «NPP» сомкнутой растительности варьирует по географической широте и водообеспеченности от 200 (субполярные области) до 2 500 г • м 2 (влажные тропические леса, вся биомасса дана в абсолютно сухом весе). Продуктивность лесов умеренной зоны около 1 ООО— 1 500 г • м 2 (см. рис. 13.39). Почти 25 % земной поверхности (около 33 млн км2) показывают годичную «N РР» — не более 500 г • м-2. Для культурных растений верхняя граница может приближаться к 7 000 г м-2 (орошаемые интенсивные культуры сахарного тростника). Культуры водорослей (например, Scenedesmus) могут в лабораторных условиях давать 10 000 г ¦ м~2 биомассы, но практическое применение их затруднено.
Глобальная «NPP» составляет по новейшим подсчетам около 210—250 млрд т биомассы или 100—120 млрд т углерода (46— 50% биомассы), из них по 50—60 млрд т углерода в океане и на суше (1 млрд т = = 1 Гт = IPg = 10IS г). На суше приблизительно половина «NPP» падает на тропики. Средняя продолжительность пребывания углерода на суше составляет 22 года, в океане — близко к неделе (планктон lt; 5 мм; значительная величина отношения поверхности к объему).
- Нетто-продукция экосистемы и биосферы
Нетто-продукция экосистемы NEP (net ecosystem production) — «продукционный параметр», который гораздо более определен, чем NPP и измеряется в большем приближении к реальности. NEP представляет собой баланс нетго-углерода в экосистеме, т.е. разницу в поглощении и выделении углерода, без рассмотрения того, где и как углерод периодически связывается в системе.
NEP более рационально получать за длительные интервалы времени (по меньшей мере за год) и на больших площадях (gt; 1 га). Базовые данные — измерение потоков С02 (внос - вынос) метеорологическими методами (измерительные вышки над гомогенным, плоским участком ландшафта). Так называемый «eddy co- variance» (вихревой ковариантный) метод измеряет с помощью трехмерного ультразвукового ветроизмерительного аппарата вертикальные брутто-потоки «квантов» воздуха и соединяет их тотчас же с открытым (open path) инфракрасным газоанализатором, измеряющим концентрации С02с очень высоким временным разрешением. Трудности этого метода измерения NEP заключаются в том, что он охватывает очень маленькую разницу в нетто очень больших потоков. Чтобы доказать, что NEP не равна нулю, требуется очень высокая точность измерений. Выбор «хороших» гомогенных насаждений ведет, как правило, к переоценке нетто- потоков на уровне всего ландшафта. Экспорт углерода в других формах, чем С02, обычным способом измерения NEP не охватывается.
Для функционирования экосистемы большое значение имеет не только поток вещества и энергии между самой системой и ее окружением, но и размещение и интенсивность потоков внутри системы. Годовой поток энергии субтропического родникового озера (рис. 13.42) несет очень небольшую фитомассу (но многочисленные генерации планктонных водорослей) и иллюстрирует, как распределяется энергия первичной продукции вдоль пищевых непей экосистемы.
В наземных биоценозах, особенно в длительно существующих лесах, круговорот вещества и энергии по отношению к фитомассе происходит существенно медленнее. Хотя в этом родниковом озере абсорбируется почти четверть излучаемой солнечной энергии, относительно густой фитопланктон может в конце концов, несмотря на отсутствие зимнего покоя, использовать для брутго-продукции только 1,2 % общего излучения. После относительно высоких потерь на дыхание (70%) для консументов и редуцентов остается только 20 %.
Чем более зрелой является экосистема, тем больше ее NEP приближается к нулю. Большое значение при таком балансе углерода имеют запасы углерода в почве (в большинстве случаев 10—20 кг - м~2). В гу- мидных тропиках они составляют часто не более 10—20 %, в бореальных хвойных лесах — 60 — 70 %, в тундре — более 90 % всего углерода экосистемы. Когда в быстрорастущих лесных посадках почвенный гумус вначале быстро разлагается (дренаж, удобрение, известкование), NEP может быть негативной, несмотря на очень позитивную «NPP». В молодых экосистемах NEP
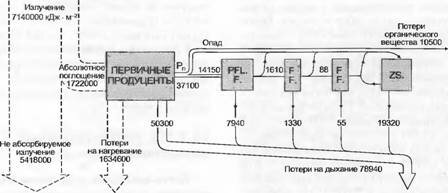
Рис. 13.42. Поток энергии через естественную экосистему планктона (субтропическое родниковое озеро Сильвер Спрингс, Флорида) (по И.Т.Odum из W. Larcher).
Значения входа и выхода даются в кДж- м'2 - а-1. Компартименты слева направо: первичные продуценты, консументы (растительноядные — PFL.F и хищники первого и второго порядков — FF.) и редуценты (ZS.)
Рис. 13.43. Глобальный круговорот углерода в среде, находящейся под воздействием человека (заштрихованы антропогенные источники С) (из Ch.Korner).
Размеры «клеток» отражают величину запасов углерода. Только около 40 % высвобождаемого ископаемого углерода остается в настоящее время в атмосфере, остаток растворяется в океане и закрепляется в наземных экосистемах (1—2 ГТС «missing carbon» — «недостающий углерод»). Эта величина примерно соответствует ежегодному высвобождению углерода из-за сведения лесов. Большие углеродные пулы в глубоких слоях океана и в известняковых породах играют роль для концентрации С02 в атмосфере только при рассмотрении очень больших периодов времени (глубинные слои океана gt; 200 лет, существенные взаимодействия с карбонатной геохимией » 1 000 лет)
большей частью положительна, в зрелых приближается к нулю, в старых деградирующих негативна. Только охват всех этих стадий развития, существующих в ландшафте (лесной области), дает информацию о том, имеет ли место закрепление в экосистеме нетто-углерода или нет.
Лес в стадии восстановления, а также спелый лес до рубки всегда имеют позитивные зі іачения NEP. Дальнейшая судьба деревьев определяет долгосрочное развитие NEP. Если в обогащенной грубым и мягким гумусом экосистеме после распада мертвой древесины вырастает следующее поколение деревьев, NEP в течение сотен лет может оставаться позитивной. Если полезная древесина перерабатывается и в конце концов рециклируется (бумага, утиль, сжигание, разложение), то калькулируемая NEP снова приближается к нулю. Деревянные постройки, например, представляют собой промежуточный фонд углерода.
Итак, объективное суждение о балансе углерода в ландшафте разрывает рамки экосистемы, поэтому для очень больших масштабов и длительных промежутков времени NEP заменяют на NBP— нетто-про- дукцию биома. NBP охватывает процессы на ландшафтном уровне, такие, как пожары, ветровалы, массовые нападения насекомых, принимает во внимание все стадии развития растительности, в том числе «окна» внутри нее, и включает также последствия любой деятельности человека. В большей части мира NBP в настоящее время негативна, это значит, что био- мы теряют нет го-углерод (корчевка лесов, интенсивная обработка почвы, экспансия густозаселенных и индустриальных территорий), хотя местами еще существуют подвергающиеся экспансии экосистемы с позитивной NBP.
Следующий шаг ведет к биосфере в целом. Ее углеродный баланс приблизительно уравновешен, так как экосистемы Земли в среднем связывают столько же углерода, сколько и отдают. Продолжающееся непрерывно сведение лесов в тропиках поставляет ежегодно в атмосферу 1 — 2 Гт углерода, еще неизученные биотические по- глошения связывают опять же 1 — 2 Гт С, причем предполагается, что это связано с удобряющим эффектом повышенного содержания С02 в атмосфере и экстенсифи- кацией природопользования в различных частях Северной Америки и Европы (вторичные и слабоиспользуемые леса). Сейчас увеличение содержания углерода в воздухе связано с тем, что человек высвобождает в атмосферу ископаемые запасы углерода в количестве 5—6 Гт, часть их растворяется в морской воде, так что ежегодно в атмосфере остается «только» почти 3 Гт дополнительного углерода. Это повышает атмосферный пул углерода (в С02) с современного значения 750 Гт в год на 0,4%, или почти I ppm (современное значение 370 ppm). Продолжение этой тенденции приведет к тому, что в конце XXI столетия общий уровень С02 по отношению к доиндустриальной эпохе удвоится (см. 13.7.6). Включенные в глобальный круговорот углерода фонды и потоки, в том числе современные антропогенные потоки, иллюстрирует рис. 13.43.
13.7.6. Биологические аспекты «проблемы С02»
В связи с тем что ископаемые источники углерода, которые большей частью начали возникать более чем 100 млн лет назад и формировались в течение многих миллионов лет, приблизительно за 200 лет (грубо говоря, с 1910 по 2100 г., когда моїуг быть исчерпаны легкодоступные запасы) в виде
О О сх
ZT
го о.
Е
amp; 320-
300 250 200 150
Тысячелетия до нашего времени
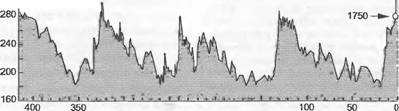
Рис. 13.44. Концентрация С02 в атмосфере за последние 0,4 млн лет на основе анализа воздушных пузырьков в антарктическом ледниковом щите (бурение на станции «Восток») (по J. Petit et al.)
360 -
С02 были выкачаны человеком в атмосферу, для растительности возникает совершенно новая ситуация. Биосфера получает новую «диету», в геологических масштабах как бы за одну ночь. Поскольку С02 — материальный базис фотосинтеза, от которого (если не считать несколько хемоавтотроф- ных видов бактерий) зависит вся жизнь на Земле, проблема С02 для экологической ботаники стала центральной. Возможность того, что обогащение атмосферы С02 также связано с климатом (так называемый парниковый эффект) и соответственно могло бы непрямым образом влиять на растения, здесь не обсуждается, речь идет только о непосредственном воздействии на растения и экосистемы.
Считается, что уровень С02 в атмосфере к началу каменноугольного периода был на уровне 2 000 — 3 000 ppm, но потом очень быстро опустился до 300 — 500 ppm, что близко к современному уровню, а также что наиболее низкие значения были в пермском периоде. Достоверно установлено, что в мелу уровень С02 сначала был очень низким (около 300 ppm), что и объясняет первое массовое распространение растений С4-типа. Механизм концентрации С02 растениями типа С4осуществляется только при очень низком уровне С02, это дает им преимущество перед растениями С3-типа. С помощью ледовых кернов из Антарктиды документирован состав атмосферы за время последних 0,4 млн лет (анализ вмороженных в лед пузырьков воздуха). Как показывает рис 13.44, концентрация С02 колебалась за это время в пределах 180— 290 ppm. Максимумы соответствуют теплым периодам, минимумы — пикам оледенений. С 1800 г., т.е. с начала отопления углем, кривая превысила этот диапазон и при-
2000 о
мерно к 1900 г. поднялась так быстро, что сейчас СОа в воздухе уже на 30 % больше, чем когда было посажено ранее большинство деревьев в парках. Почти все сейчас существующие виды растений появились в свободных ото льда областях в тот период, когда уровень С02 был равен 180 ppm (иначе бы они просто вымерли; в последний раз концентрация С02 была столь низкой приблизительно 20 000 лет назад). Около 1990 г. они пережили удвоение этого значения (в 2000 г. — 370 ppm). Если не произойдет глобального коллапса мирового хозяйства, концентрация С02 за последующие 100 лет еще удвоится.
Исходное положение всех рассуждений о биологических последствиях повышения С02 лежит в кривой зависимости нетго-фотосин- теза от концентраций С02 (рис. 13.45, см. 6.5.11.2). Она показывает, что С3-растения могут существовать при еще больших концентрациях, чем сейчас, и больше фотосинтезировать. Разумеется, такие кривые являются как бы «снимками данного момента», которые только доказывают, что при обеспечении RubisCo в данный момент и без ограничения отложения ассимилятов процесс карбоксилирования еще не насыщен С02. Как было изложено в разделах 13.7.2 и 13.7.3, ускорение роста зависит еще от многих других факторов, и только в тех случаях, когда не существует ограничения отложения, концентрация С02 может быть причислена к стимуляторам роста.
Прирост урожая в тепличном садоводстве равняется примерно 30 % в сезон, если растениям предоставляется уровень СО, 600 ppm или более, что уже перед Второй мировой войной использовалось в немецких и голландских теплицах. Лучше всего удобренные и орошенные пшеничные поля в Аризоне давали на 14 % больше зерна, когда культивировались при уровне С02 около 600 ppm. Такие цифры должны хорошо смотреться на фоне того, что урожаи пшеницы благодаря новым сортам и оптимальному уходу в последние 100 лет уже увеличились на 300-500%.
Когда все другие ресурсы, кроме С02, относятся к ресурсам, лимитирующим рост, что в естественных условиях происходит почти всегда, остаются три возможности для долгосрочного реагирования:
• редукция емкости фотосинтеза (меньше RubisCo и тем самым меньше азота на листовую поверхность, или меньше листьев, т.е. более низкая LAR, см. 13.7.3);
- более высокий экспорт углерода (например, более быстрый обмен тонких корней, экссудация корней, экспорт в микоризу, выделение изопренов);
- усиление роста с помощью разбавления питательных вешеств, особенно N, т. е. продуцирование биомассы с более высоким отношением C/N.
Все три пути идут, как правило, параллельно. Обычно происходит сокращение емкости фотосинтеза, но очень редко полностью, так что при повышенном содержании С02 на единицу листовой поверхности почти всегда связывается больше углерода (рис. 13.45).
Многочисленные эксперименты показывают, что экспорт углерода при повышенном С02 усиливается. Наблюдалось, что повышенное содержание растворимых углеводов в ризосфере ведет к усилению связывания азота почвенными микроорганизмами, следствием чего могут быть симптомы азотного голодания. Увеличение соотношения C/N наблюдалось почти всегда и прежде всего в листьях. В большинстве случаев общее содержание структурно не связанных углеводов повышается (например, крахмал, сахар; англ. non structural carbohydrates— NSC). Со снижением содержания протеина (N) и увеличением содер-
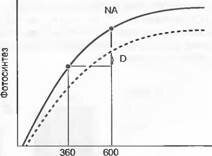
Концентрация С02, ppm
Рис. 13.45. Регулирование зависимости фотосинтеза от концентрации С02 при длительном воздействии повышающегося содержания С02 (down regulation — пунктирная кривая) зависит от условий роста (активность поглощения ассимилятов) и возраста растений.
Точка NA отмечает увеличение нетто фотосинтеза при повышении концентрации С02 до 600 ppm без подобного регулирования; D — сохранившаяся нетто-прибыль после регулирования
О -1,0
’ 0
10 1 5 20 25 30 35 40
Возраст деревьев, годы
Рис. 13.46. Воздействие повышенного содержания С02 на длительный рост деревьев в окрестностях геологического источника С02 в Тоскане (Раполано) (по S.Hattenschwiler et al.).
1000L ? 360 ppm С02 Прерия в Канзасе
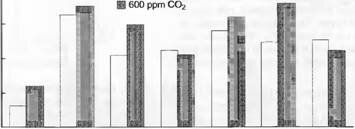
1989 1990 1991 1992 1993 1994 1995
Сухая Нормальная Сухая Сырая Сырая Сухая Нормальная
Рис. 13.47. Действие повышенного содержания С02 на участке высокозлаковой прерии. Только в сухие годы повышение концентрации С02стимулирует накопление биомассы, это результат воздействия С02 на устьица, транспирацию и тем самым на влажность почвы (600 вместо 360 ppm в больших «open top» («открытых сверху») камерах) (по С.Owensby et al.)
На примере анализа годичных колец каменного дуба (Quercus ilex) можно увидеть, что деревья на ранних фазах развития близ источника по сравнению с более удаленными контрольными деревьями росли явно быстрее (рис. показывает разницу в приросте по отношению к деревьям в стороне от источника). Примерно в 30-летнем возрасте сигнал исчезает (индекс «опыт»/ контроль нулевой; сплошная линия). Полосы дисперсии показывают среднюю вариабельность в каждом случае для 10 деревьев. С помощью углеродного (14С) анализа древесины годичных колец установлено, что подвергавшиеся воздействию источника деревья фактически всегда испытывали вдвое более высокую концентрацию С02, чем контрольные (геологический С02 свободен от 14С, путем смешивания его с нормальным воздухом может быть реконструирована средняя концентрация С02, которую испытывали деревья) жания NSC (С) изменяется и качество питания. Для травоядных животных доказано, что это отрицательно влияет на их рост и размножение.
-и.ь -
Quercus ilex
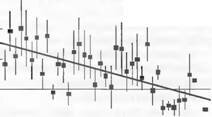
у - 0,791 - 0.024х г2 = 0,46 Рlt; 0,001
Прирост биомассы в естественной растительности (почти все данные по злаковым сообществам) обычно очень незначителен (менее 15 % при смоделированном удвоении концентрации С02) или даже равен нулю. Для естественных лесов (85 % глобального запаса биомассы) данных не имеется, по реакция каменных дубов, растущих на границе с естественным источником С02в Тоскане, дает первые доказательства возможной реакции: стимуляции роста на самой ранней ювенильной фазе развития при удовлетворительном предоставлении света и питательных веществ. С увеличением возраста реагирование становится все меньшим до того момента, когда эффект сводится к нулю (рис. 13.46) Это могло бы означать некоторое убыстрение жизненного цикла. Более высокий запас биомассы на единицу земной поверхности с этим неизбежно не связан. Немалое число С02-экспериментов со всходами деревьев дало доказательства этого только на очень ранних стадиях. Большинство видов реагирует на С02 положительно, но очень видоспецифично, причем различия зависят от синхронного снабжения светом и питательными веществами, поэтому однозначные выводы невозможны.
При всех реакциях растений на С02 было установлено, что различные виды откликаются очень по-разному. В отдельных случаях даже і габлюдалось, что различные генотипы одного вида реагируют не одинаково. Это означает, что проблема С02 в любом случае может рассматриваться как проблема биоразнообразия особого рода. Снабжение С02 влияет на конкурентные отношения между видами. Можно согласиться с тем, что этот фактор влияет на структуру видового состава в мировом масштабе.
Кроме прямого воздействия С02 на фотосинтез и последующее употребление ассимилятов имеется еще и непрямое влияние С02 через водный режим. Устьица часто реагируют на повышенную концентрацию С02 сужением устьичной щели (см.
Ежегодной выработке ископаемого углерода противопоставляется содержание его во всей вырубаемой национальным лесным хозяйством древесине или, как альтернатива, масса углерода, получаемая с плантаций биомассы (например, китайского тростника) на 10% современных посевных площадей
Рис. 13.48. Потенциал замещения ископаемого углерода биологическими источниками углерода (по Ch.Korner).
- . Реакция зависит от влажности воздуха и почвы, не у всех видов она одинаково сильная, а у деревьев более выражена на стадии проростков, чем у взрослых экземпляров. В злаковых сообществах эта реакция устьиц вызывает более медленное снижение влажности почвы в периоды, бедные осадками, что является причиной увеличения аккумуляции биомассы. Большая часть тех явлений, которые происходят в природной злаковой растительности, могут сводиться к этому непрямому эффекту. В сухие годы данный эффект соответственно проявляется сильнее, чем ВС влажные (рис. 13.47). Предполагается, чтс эффект замедления снижения влажности е насыщенном С02 пространстве станет реальностью, если условие испарения по сравнению с современными не изменяется.
Антропогенные выбросы С02 в настоя шее время составляют почти 6 Гт С-а-1, и будут увеличиваться далее, так что растительность в течение следующих 100 или более лет не сможет связать поступающий излишек. Гораздо вероятнее, что наземное использование дополнительного углерода переместится в атмосферу. Уже сейчас, еще на крутой ветви кривой функции зависимости фотосинтеза от С02 при превышении С02 на 30 % по сравнению с доинду- стриальным временем максимальное нет- то-связывание составляет только 2 Гт (даже если расчетное поглощение наземного углерода (см. рис. 13.43) приписать исключительно удобряющему эффекту С02, что, вероятно, переоценивается) — это удивительно малая часть наземной первичной нетго-продукции, составляющей около 60 Гт С • а Вследствие стехиометрического связывания Р и N и других ограничений относительный эффект увеличения концентрации С02 значительно уменьшается.
Био-менеджмент усиления связывания углерода растениями рационален и в экологическом отношении полезен, но при этом не следует питать иллюзий, что абсолютные значения его взаимосвязаны с эмиссией С. Только существенное расширение лесных площадей могло бы долговременно способствовать связыванию углерода в биомассе. Но в данный момент лесные площади массированно сокращаются. Облесение выкорчеванных площадей постепенно возмещает потери, но не дает нетто- прироста, пока они не восстановятся до предыдущего состояния, а это происходит со 100 — 200-летней задержкой. Старые деревья содержат существенно больше С, чем молодые, поэтому замена старых насаждений молодыми всегда (!) представляет собой потерю углерода. Следует помнить о том, что в долговременном балансе углерода решающим является не оборот, а- величина общих фондов (средняя продолжительность пребывания С в ландшафте). Быстрорастущие лесные посадки не вносят вклад в восполнение углеродного «долга» местности, хотя и могут служить ценным источником обновляемых ресурсов (замена ископаемым источникам углерода). Чтобы порядок величин замещающего потенциала был очевидным, на рис. 13.48 показано, сколько ископаемого углерода теоретически могло быть замещено биомассой, если бы для этого была введена в действие вся (!) вырубаемая за год древесина в Германии, Австрии или Швейцарии (другое применение древесины и техническое использование заменителей не предполагаются). Цифры свидетельствуют, что проблема не разрешима ни таким образом, ни созданием специальных плантаций биомассы, или биомасса редуцируется до немногих процентов от ископаемого углерода. Это не означает, что использование биомассы вместо ископаемого углерода не приветствуется и не приносит экологических и хозяйственных выгод. Никакой эффект биомассы не может хотя бы примерно приблизиться к потенциалу восстановления даже при маленьких (менее 5 %) квотах сбережения при потреблении горючих веществ, причем технически возможная экономия вместе с той, которая вытекает из действий человека, без сколько-либо значимого снижения уровня жизни может превышать 50 %. Не следует таким образом упускать из вида порядок величин, если в решении проблемы рассчитывать на роль растений.
- Биотические взаимодействия
Биоценозы Земли характеризуются не только основополагающими пищевыми цепями от продуцентов к консументам и редуцентам, но также и многими другими аспектами совместного существования и конкуренции организмов. Биотические взаимодействия, обычно называемые интерференцией, могут проявляться между индивидами одной популяции (вида) или между индивидами разных видов. Между экологически самостоятельными видами они обычно еще очень слабые, но могут углубляться вплоть до полной взаимозависимости (например, при симбиозе или паразитизме). Для популяционной биологии или популяционной генетики они имеют основополагающее значение. Ниже будут сопоставлены некоторые важные биотические взаимодействия; решающим критерием при этом будут позитивные (+), негативные (-) влияния на интенсивность возобновления или их отсутствие (0), которые оказывают друг на друга два партнера (А и В);
|
А gt; В |
В gt; А |
Тип взаимодействия |
|
- |
- |
Конкуренция |
|
+ |
- |
Паразитизм, поедание |
|
+ |
+ |
Кооперация, симбиоз |
|
+ |
0 |
Комменсализм |
|
0 |
0 |
Нейтрализм |
Биотические взаимоотношения между автотрофними растениями простираются от конкуренции до кооперации, причем пространственное вытеснение и борьба за свет и питательные вещества, равно как и за воду в почве, играют роль в такой же степени, как и изменения климата биоценоза (см.
- или химические воздействия (алле- лопатия, см. 9.5) и др. Некоторые из этих комплексных взаимоотношений можно понять на основе опытов в культуре с двумя или несколькими видами (рис. 13.49), другие же настолько сильно зависят от широкого комплекса влияний внешней среды, что в поле их можно разграничить только приблизительно путем наблюдений (пространственно-временные модели) или с помощью манипулятивного вмешательства.
Например, в то время как ряска Spirodela (Lemna) polyrhiza всегда размножается интенсивнее, чем Lemna gibba, последняя в плотной смешанной культуре подавляет Spirodela в ходе конкуренции за свет (рис. 13.49). Насколько сильна корневая конкуренция за питательные вещества, показывает опыт, при котором молодые растения ели в березовом лесу удобряли маркированными фосфорными соединениями. Если березовые корни обрезают, то ели могут усваивать в 5 — 9 раз больше фосфора. Такие («zoot- trenching» — траншейно-корневые) опыты в тропическом лесу также документируют мощную корневую конкуренцию между всходами и взрослыми растениями.
При культивировании важнейших видов на газоне из Lolium регеппе высокорослый Trifolium pratense дает более высокую продукцию биомассы, чем низкорослый Т. repens. Все виды вместе через год почти удваивают продукцию, но если удаляют «мешающий» им Lolium регеппе, и в целом становятся в три раза более продуктивными, чем один Т. pratense. Здесь очевидно явление кооперации, или комменсализма: связывание азота воздуха видами клевера с помощью клубеньковых бактерий явно способствует успеху других видов. Шпалерный кустарничек Loiseleuria (см. рис. 12.9,Е) в аркто-альпийских обдуваемых ветрами местах часто существует совместно с кустистыми лишайниками (Cetraria и др.), которым он предоставляет условия для закрепления; в свою очередь возвышающиеся над ковром листьев талломы лишайников тормозят ветер, что создает для Loiseleuria более благоприятные микроклиматические условия.
Всходы и ювенильные растения обычно намного сильнее страдают от конкурентной борьбы, чем устоявшиеся взрослые растения. У лиственницы и сосны многие взошедшие семена выживают на примитивных почвах или на оголенных местах и, напротив, почти не выживают среди яруса высоких трав, мхов или кустарничков, которые разрастаются под более старыми деревьями. Здесь и световые условия неудовлетворительны, и влажность воздуха выше, одревеснение проростков меньше и поэтому значительно сильнее поражение грибами и поедание улитками. Так же и многие другие пионерные древесные растения препятствуют возобновлению лиственницы и сосны и способствуют сукцессии другими видами деревьев.
При аллелопатическом ограничении (см.
- подроста особей других видов (но частично и своего вида) значение имеют продукты обмена веществ. Примеры этому найдены у водорослей Chlorella и Nizschia, у богатых терпеноидами Lamiaceae (например, зоны торможения вокруг видов Salvia в Калифорнии) и Myrtaceae (например, почти свободные от подлеска посадки Eucalyptus). Бросается в глаза «враждебность» к подлеску также у Robinia, Juglans и многих хвойных деревьев.
К отношениям между гетеротрофами и растениями относятся аллелопатия продуцирующих антибиотики актиномицетов и грибов против бактерий и обнаруженное К. Клеем с сотрудниками защитное действие эндофитных грибов в ткани листа против фитофагов. Существенны взаимодействия автотрофных растений с грибными симбионтами в лишайниках (см. 9.2.4) и микоризе (см. 9.2.3), а также с комменсалами, например с многочисленными сапрофитными бактериями и грибами, которые живут на опадающих частях растений, или с паразитами (многие
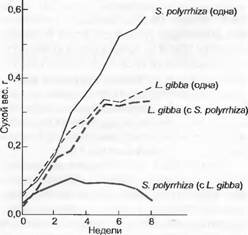
Рис. 13.49. Размножение двух свободноплавающих видов Lemnaceae (Spirodela polyrhiza и Lemna gibba) в чистой культуре и смешанной культуре в условиях конкуренции (по I. Harper)
бактерии и грибы, некоторые покрытосеменные; см. 9.1.1, бокс 11.4).
Значительные изменения растительности могут возникать, когда определенные виды деревьев сильно поражаются или полностью элиминируются грибными заболеваниями; за последние десятилетия, например, это касается полевого вяза в Европе (Ulmus minor; гриб — Ophiostoma ulmii, см. 11.2) и восточно-североамериканского каштана (Castanea dentata\ изначально поражается до 60 % частей дерева; гриб — Endothia parasitica, занесенный в 1904 г. из Китая; см. Diaporthales). Остается лишь ожидать, что произошел отбор резистентных биотипов этих деревьев, которые могли бы вновь завоевать утерянные видом территории.
Особенно многосторонни и экологически значимы биотические взаимоотношения между растениями и животными. В первую очередь здесь следует назвать фитофагов, т.е. травоядных животных, как консументов первого порядка. При этом причиной очень значительных повреждений и изменений растительного покрова могут стать насекомые (тли, короеды, пяденицы), улитки или млекопитающие (мелкие грызуны, кролики, жвачные животные), поедающие или высасывающие вегетативные органы, цветы и во многих случаях семена. Обусловленное этим «давление» отбора приводит к образованию многочисленных защитных механизмов: колючки, шипы, жгучие волоски, иглы кристаллов, горькие и ядовитые вещества и т.д. (см. 4.2.6,
- 11.168). Особый случай — образова ние животными галлов (см. 9.1.1). Симбиотические связи с животными проявляются у семенных (а также и у низших) растений особенно в области биологии цветения, плодоношения и семеношения (см. бокс 11.4). Многие бактерии и грибы паразитируют на животных. Немногие грибы и покрытосеменные специализировались как «ловцы животных» (см. 9.1.2).
Пастбищные животные при стравливании повреждают в первую очередь молодые проростки древесных растений и благоприятствуют тем самым хорошо регенерирующим злакам и крупным травам лугов. Дальнейшие изменения местообитаний происходят путем вытаптывания (уплотнение почвы, механические повреждения) и удобрения. Как следствие этого распространяются растения — так называемые «вредители пастбищ», которые скот обходит, например в Центральной Европе Juni- perus communis, Berberis vulgaris, Prunus spino- sa, Ononis spinosa, Eryngium campestre, Cardu- us sp., Cirsium sp., Nardus stricta с твердыми и колючими побегами, а также виды Ru- тех, Ranunculus, Euphorbia, Apiaceae, Lami- aceae, Liiiales (например, Colchicum autum- nale), в них содержатся горькие, ароматические или ядовитые вещества.
История развития многих родственных групп покрытосеменных была успешной, а их формообразование богатым, очевидно, потому, что они выработали действенные химические вещества, защищающие от поедания животными, например Capparales с их гликозидами горчичного масла, многие Gentianales с индол-алкалоидами или Sola- пасеае с тропан-алкалоидами (см. 9.4.1). Только определенные группы фитофагов могут потреблять эти защитные вещества без вреда для себя и даже специализируются на соответствующих растениях (например, бабочки Pierinae на Capparales). Бабочка-монарх (Danaus plexippus) встраивает получаемый из своих кормовых растений (Asclepiadaceae) ядовитый гарденолид-тикозид даже в тело гусеницы и взрослого животного и поэтому несъедобна для своих врагов.
В качестве примера еще недостаточно исследованных взаимодействий между растениями и муравьями (см. зоохория) можно привести неотропические виды Acacia (например, А. сог- nigera). У этого дерева из влажных тропических лесов развит симбиоз с агрессивными муравьями (Pseudomyrmex fermginea): оно служит им жилым помещением, источником питания и эк- страфлорального нектара (см. рис. 11.251) и при этом очень успешно «использует» муравьев, как защиту от всех фитофагов. Муравьи даже подрезают и удаляют разрастающиеся лианы и кон • курирующие соседние растения, чтобы их растение-хозяин могло лучше развиваться. Эффективность этого симбиоза видна из сравнения с другими акациями, которые не заселяются муравьями: они сильно подавлены в росте и искривлены. Аналогичные отношения имеются между муравьями и пионерными деревьями рода Cecropia.
Очень существенна для многих растений потеря семян из-за животных. Fagus sylvatica может успешно размножаться только в урожайные годы с повышенной семенной продуктивностью. Такие годы повторяются через неодинаковые промежутки времени; это следствие того, что паразитирующие в семенах насекомые в своих циклах развития не могут ориентироваться только на урожайные годы. Неотропические бобовые выработали две защитные стратегии против жуков (Bmchidae), поедающих их семена: либо они продуцируют неядовитые, но многочисленные и мелкие семена, из которых хотя бы часть остается нетронутой, либо образуют более крупные и немногочисленные семена, но содержащие ядовитые вещества (см. 11.2, Fabales).
Все эти положительные и отрицательные взаимодействия влияют на рост популяций, участвующих в создании биоценоза. Одни виды становятся доминирующими, другие остаются подчиненными или исчезают, что свидетельствует о лабильном или более или менее стабильном состоянии равновесия. Такие примеры можно описать математически и симулировать с помощью компьютерных моделей.
Конкурентная борьба между двумя видами становится тем острее, чем ближе их экологические потребности. Надолго их совместное существование в одной и той же экологической нише невозможно (см. 12.1). Поэтому при негативных биотических взаимодействиях мы постоянно находим, что существующие в сообществе виды «уклоняются» от конкуренции: в пределах генетически установившейся в чистой культуре широты нормы реакции видов виды «сдвигаются» в ту или иную сторону при произрастании в смешанной культуре, т.е. под влиянием биотических взаимодействий их распространение таково, что перекрывание с амплитудами и областями оптимума конкурентов оказывается минимальным.
Этот принцип уже обсуждался в разделе 12.1. Он применим, например, и к важным луговым злакам Центральной Европы: по отношению к высоте уровня грунтовых вод (влажности почвы) в чистой культуре они обнаруживают очень близкие оптимумы роста, в то время как в смешанной культуре оптимумы роста отстоят друг от друга достаточно далеко. Bromus erectus и многие другие «ксерофильные» виды в действительности очевидно, не «сухолюбивые», а только более «суховыносливые». Также и Oxalis acetosella не «тенелюбива», а «теневынослива». Многие реликтовые средиземноморские виды лишь потому ограничиваются в своем распространении недоступными трещинами скал, что во всех' других местообитаниях они стравливаются козами и овцами.
Экологическая позиция и широкая или соответственно узкая амплитуда экологических потребностей видов (эвритопные, соотв., стенотопные) очень зависят также от их биоценотических партнеров, она относительна. Во всяком случае эти биотические взаимоотношения между видами очень разнообразны, комплексны и системно объединены друг с другом и прочими факторами местообитания; это решающим образом способствует стабильности и саморегуляции экосистем.
Виды, которые в своих исконных экосистемах занимают подчиненное положение и «находятся под контролем», могут в «чужих» экосистемах становиться агрессивными «сорняками», поскольку там отсутствуют их естественные враги. Примерно так произошло с европейско-атлантическим Ulex europaeus в Новой Зеландии, европейским Hypericum perforatum в Северной Америке или неотропической Opuntia inermis в Австралии. Только намеренный занос питающейся Opuntia венесуэльской моли Cacto- blastis cactorum ликвидировал этот вредный сорняк на площади более 120 млн га в течение нескольких лет. Похожая ситуация с водяной чумой Elodea canadensis в Европе. Напротив, интродуцированные животные (например, козы и кролики) в значительной степени уничтожили малоустойчивые к выпасанию островные флоры, например Еавайских островов, о-ва Св. Елены. Многие виды биоценоза играют несколько экологических ролей, так, Viscutn album (омела) для деревьев-хозяев — полупаразит, для разносящих семена птиц — симбионт, для различных насекомых-фитофагов — даже хозяин.
Давление паразитов и других врагов на вид тем сильнее, чем больше этот вид стремится к доминированию и образует большие сомкнутые популяции. Например, гомогенные искусственные еловые посадки намного более восприимчивы к носящим характер эпидемий вспышкам численности вредителей (короедов, пядениц), чем естественные смешанные насаждения ели с другими древесными породами. Богатое видовое разнообразие многих лесов от умеренно-теплых до тропических, видимо, объясняется тем, что каждый древесный вид, разрастающийся за счет другого, вскоре вновь редуцируется из-за богатой паразитической фауны и флоры.
- Использование биомассы и земель человеком
Таблица 13.5. Мировые урожаи продуктов растительного происхождения, употребляемых человеком (ФАО, 1999)
|
Вид продукта |
Сырой вес, 106 т |
Вид продукта |
Сырой вес, 106 т |
|
Хлебные злаки |
2064 |
Сахаристые растения |
1538 |
|
Кукуруза |
600 |
Сахарный тростник |
1 275 |
|
Рис |
596 |
Сахарная свекла |
263 |
|
Пшеница |
584 |
||
|
из них сахара-сырца |
133 |
||
|
Ячмень |
130 |
||
|
Зернобобовые |
59 |
||
|
Просо |
89 |
||
|
Фасоль |
19 |
||
|
Овес |
25 |
||
|
Горох |
12 |
||
|
Рожь |
20 |
||
|
Прочие (гречиха и др.) |
19 |
Нут |
9 |
|
Крахмалоносные клубни |
650 |
Бобы |
4 |
|
и корни |
Прочие (чечевица и др.) |
15 |
|
|
Картофель |
294 |
Масличные и жировые культуры |
483 |
|
Маниок |
168 |
(плоды, семена) |
|
|
Батат |
135 |
Соевые бобы |
154 |
|
Прочие (ямс, таро и др.) |
52 |
Плоды масличной пальмы |
98 |
Население мира развивалось скачкообразно: с 10 млн человек 10000 лет назад оно выросло за 2 000 лет примерно на 160 млн, достигнув к 1850 г. почти 1,2 млрд человек, к 1988 г. — 5 млрд и в 2000 г. — уже 6 млрд человек. Человечество и домашние животные в отношении биомассы представляют собой огромную величину: около 100 млн т людей и 400 млн т домашних животных по сравнению с остальной наземной биомассой животных, составляющей 2 300 млн т (в сухом весе). Населению Земли для питания необходимо ежегодно 1 200 млн т зерна и других растительных продуктов. Сырье для этого со всеми потерями соответствует почти 10 % всей наземной первичной продукции (120 млрд т в год) с примерно 10% земной поверхности. На питание животных расходуется ежегодно около 130 млн т сельскохозяйственной и около 36 млн т рыбной продукции. Общая годовая хозяйственная потребность в древесине соответствует фитомассе 2,7 млрд т.
Эти немногие цифры иллюстрируют, что питание человека в чрезвычайной степени влияет на растительный покров Земли. В отношении своего питания человек полностью зависит от использования зеленых растений, непосредственно использует пищевые растения (например, хлебные злаки, плоды бобовых, крахмалосодержащие клубни, сахарную свеклу и сахарный тростник, плоды масличных растений, фрукты и овощи), опосредованно (через растительные кормовые источники) — разводимых животных (например, рыбу или млекопитающих), дающих мясо, жир и молочные продукты. Возбуждающие продукты и лекарственные средства (например, вино, пиво, кофе, табак, антибиотики, сердечные гликозиды, алкалоиды), промышленное сырье (например, древесина, волокнистые материалы, каучук) и источники энергии (дрова, уголь, нефть) добываются также большей частью из растительного царства.
Окончание табл. 13.5
|
Вид продукта |
Сырой вес, 106т |
Вид продукта |
Сырой вес, 106 т |
|
Семена хлопка |
52 |
Прочие (сливы, папайя, финики, |
71 |
|
Кокосовые орехи |
47 |
земляника, абрикосы, вишни, авокадо и мн. др.) |
|
|
Рапс |
43 |
Орехоплодные (миндаль, грецкие |
7 |
|
Арахис |
33 |
орехи, кешью, лесной орех, |
|
|
Подсолнечник Маслины Прочие (семена льна, кунжута и др.) |
28 13 13 |
настоящие каштаны и мн. др.) Вкусовые продукты Вино Табак |
47 28 7 |
|
Овощи |
559 |
Кофе |
6 |
|
Томаты |
95 |
Какао |
3 |
|
Капуста |
49 |
Чай |
3 |
|
Лук |
44 |
Пряности |
5 |
|
Огурцы |
29 |
Растительные волокна |
24 |
|
Баклажаны |
21 |
Хлопок |
18 |
|
Морковь |
18 |
Джут |
3 |
|
Перец сладкий Прочие (салат, тыква, цветная |
18 284 |
Прочие (лен, сизаль, конопля и др.) Каучук |
3 |
|
капуста, сладкая кукуруза, шпинат |
7 |
||
|
и др.) |
Животные продукты |
846 |
|
|
Фрукты |
515 |
Мясо |
226 |
|
Цитрусовые |
98 |
Молоко |
562 |
|
Бананы (фруктовые и кулинарные) |
89 |
Яйца |
54 |
|
Виноград |
61 |
Мед |
1 |
|
Яблоки |
60 |
Овечья шерсть |
ок. 2 |
|
Арбузы |
52 |
Кормовые культуры |
5083 |
|
Манго |
24 |
Люцерна |
521 |
|
Дыни |
19 |
Кукуруза |
472 |
|
Груши |
16 |
Прочие (злаки, клевер) |
529 |
|
Ананасы |
13 |
Кормовые свекла, тыква и др. |
1 645 |
|
Персики и нектарины |
12 |
Сено |
1918 |
Примечание. Выделенные жирным шрифтом суммы из-за округления последнего знака не точно соответствуют составляющим суммы числам.
Рассматривая абсолютные величины пищевой продукции (табл. 13.5), можно удивляться тому, что сам человек в сущности живет только за счет немногих представителей одного семейства растений, именно злаков (Роасеае), среди которых только четыре изначально специфичные для своих континентов — рис, пшеница, кукуруза, просо — представляют собой главные пищевые растения. С учетом пастбищных и кормовых злаков, служащих для питания крупного рогатого скота, овец и коз, эти злаки являются и косвенной основой нашего питания. Все другие культурные растения вместе по своему объему (в сухом весе) отстоят далеко.
- Использование и преобразование растительности
¦-Прочие земли
Коренные леса Вторичные леса и лесные посадки ^-Естественные злаковые сообщества -Пастбища Площади - под растениеводством 1300 1500 1700 1850 2000

Рис. 13.50. Изменение площадей от близких к естественным экосистем (особенно коренные леса, естественные злаковые сообщества), до экстенсивно используемых экосистем (особенно вторичные леса, лесные посадки, пастбищные земли) и интенсивно используемых экосистем (растениеводство) со средних веков до современности. Разрушение лесов происходит намного быстрее, чем развитие новых культурных ландшафтов (по P. Buringh и R. Dudal)
Использование человеком земель рассматривается в настоящее время как наиболее значимое воздействие на планету, еще более радикальное значение имеет это для будущих поколений, так же как, например, неоднократно обсуждавшиеся изменения в атмосфере (см. 13.7.6). Этот процесс идет параллельно с биологическим обеднением (потерей биоразнообразия) и истощением почв (потерей плодородия). Как исчезновение видов, так и потеря плодородных почв в исторически обозримых рамках необратимы. Современное обедне ние биосферы сравнимо в геологических масштабах времени с последствиями метеоритных ударов, которые в истории Земли могли бы завершать крупные эпохи эволюции (например, время динозавров). Где и каким образом человечество добывает пищевые и кормовые растения и другое растительное сырье, сегодня и в будущем накладывает отпечаток на состояние и функции биосферы. Разделение на природные и хозяйственные ландшафты сейчас все более затруднительно, поскольку даже на первый взгляд еще естественный, привлекательный ландшафт часто очень незначительно уже антропогенно изменен. Основными параметрами изменения являются:
- изъятие биомассы — выборочное или сплошное (сукцессионное изменение экосистем);
- преобразование экосистем: лес (корчевание) —gt; саванна, пастбище;
- инвазия экзотических растений, животных и микроорганизмов;
- дистанционное воздействие загрязняющих воздух веществ, перекармливание (С02, NO„), изменение климата;
- планомерное разведение полезных растений (полеводство, лесные плантации);
- замена биосферы антропосферой с сильно «опечатанной» почвой (поселения, транспортные пути, индустриальные земли).
Условие для выживания человечества — использование и организация биосферы, ориентированные на экологическую устойчивость (sustainability), а не на эксплуатацию (exploitation) и неограниченный рост, при особом внимании к охране природы и окружающей среды. Рис. 13.50 показывает, каких опустошительных масштабов достигло уничтожение лесов (особенно в тропиках) за последние 40 лет; в сравнении с этим создание новых культурных ландшафтов относительно отстает. Так, близкие к естественным или естественные биоценозы в плотнонаселенных областях Земли (например, в Центральной Гвропе) сократились до крошечных пятен или совсем исчезли. Поэтому реконструкция потенциальных экосистем (даже при абстрагировании их от воздействия человека) сейчас на больших территориях едва ли возможна.
Еще по теме Продукция биомассы:
- Биомасса, продуктивность, глобальный круговорот углерода Запасы биомассы
- БИОМАССА И ПОТОК ЭНЕРГИИ
- ОСОБЕННОСТИ ХИМИЧЕСКОГО СОСТАВАЖИВОГО ВЕЩЕСТВА И БИОМАССЫ
- Первичная продукция.
- Исследование процессавыделения биомассы микроорганизмовиз культуральной жидкости
- САМАЯ БОЛЬШАЯ БИОМАССА ПЛАНЕТЫ
- СОКРАЩЕНИЯ ПЛОЩАДИ И БИОМАССЫ ЛЕСОВ
- Продукция
- ЗАПАСЫ И ПРОДУКЦИЯ ДРЕВЕСНОГО ЯРУСАКИРСАНОВСКОГО БОЛОТА Е. А. Головацкая, М. В. Волознева, Е. В. Порохина
- Размеры заготовок и продукция сайгачьего промысла
- Состав пчелиной семьи и ее продукция
- Определения содержания нитратов в растительной продукции.
- Растительные токсины, влияющие на технологические свойства и биологическую ценность животноводческой продукции
- К МЕТОДИКЕ ОПРЕДЕЛЕНИЯ ЛИНЕЙНОГО ПРИРОСТА И ПРОДУКЦИИ СФАГНОВЫХ МХОВ НА МЕЗО-ОЛИГОТРОФНЫХБОЛОТАХ ЗАПАДНОЙ СИБИРИ
- О влиянии свойств почв и удобрений на качество растительной продукции.