Самоподдержание и популяциях растений
Важной характеристикой особей растений является процесс их репродукции, или размножения, обеспечивающий их самоподдержание в популяцнх (Левина, 1967, Злобив, 1992, Злобин, 1993).
Репродукция является ключевым звеном в динамике популяций я определяет их устойчивое существование в экосистемах.В зависимости от механизма образования диаспор различают три основные формы репродукции:
- половая репродукция, или генеративное размножение, связанное с образованием гамет, их копуляцией и формированием зиготы, обладающей комбинированным генотипом;
- бесполое размножение, состоящее в образовании спор; как способ размножения свойственно споровым и низшим растениям;
- вегетативное размножение, осуществляемое за счет специализированных структур, несущих почки, или за счет партикуляции материнских особей.
Репродукция - многоэтапный процесс, контролируемый как состоянием особей, так и популяционными параметрами. Вследствие этого существует большое количество показателей и оценок уровня репродукции.
- Половая репродукция
Половая репродукция у покрытосеменных растений состоит в последовательном образовании цветков, плодов и семян и является основным способом их размножения. Этой проблеме посвящена большая биологическая и геоботаническая литература (Fryxell, 1957; Persival, 1965; Левина, 1981; Willson, 1983; Fenner, 1985: Злобин, 1989 и др.).
При оценке полового размножения растений с точки зрения их популяционной жизни одним из важных показателей является радиус репродуктивной активности, который определяет расстояние эффективного переноса пыльцы и зачатков размножения (спор, семян, плодов). Это пока мало изученный параметр, при оценке которого приходится дополнительно учитывать фертильность пыльцы и жизнеспособность семян.
Репродуктивный процесс контролируется на двух уровнях - на уровне отдельных особей и на уровне популяций.
Действительно, первые фазы репродукции (органогенез цветков) полностью зависят от состояния особей, тогда как завершающие фазы этого процесса - опыление и распространение семян - требуют присутствия партнеров и обусловлены структурой популяции и ее экосистем- ными связями. Косвенно, через изменение жизненного состояния растений популяционные особенности воздействуют и на первые фазы репродукции. Репродуктивные стратегии разных популяций одного и того же таксономического вида могут различаться.Уровень особей. Состояние и функционирование генетов и раметов являются основными факторами успешности полового размножения. Например, у душистого колоска Anthoxanthum odoratum вес семян на 17% определялся генетическими особенностями материнской особи, на 3% - особенностями отцовской формы и на 80% зависит от экологических условий произрастания материнского растения (Antonovics, Schmitt, 1986).
Особенности семян определяются жизненной формой растений. В ряду «деревья - кустарники - травы* происходит уменьшение размера плодов и семян примерно в 10 раз на каждый переход (Silvertown, 1982). Зависимость всхожести семян от их размера не установлена. В одних случаях лучше прорастают крупные семена, в других - мелкие. Но проростки из более крупных семян всегда имеют повышенную жизнеспособность и лучшую выживаемость.
Не следует упускать из виду, что мелкие и слаборазвитые диаспоры - это важный резерв популяции. Как показано К.Г. Ткаченко (2006), всходы из крупных семян высокой жизнеспособности чаще гибнут от поздних весенних заморозков, и популяция выживает за счет второй волны всходов из мелких диаспор с малым запасом питательных веществ.
Основным показателем репродуктивного потенциала растений является количество производимых семян в расчете на особь. Принято различать две основные категории семенной продуктивности высших растений:
а) потенциальную семенную продуктивность, определяемую количеством закладываемых на одной особи семязачатков;
б) реальную семенную продуктивность, понимаемую как количество зрелых полноценных семян, продуцируемых одной особью.
Формируемые на растениях семена имеют разный уровень жизнеспособности. Г.Г. Жиляев (2003) в специальном исследований на большой группе травянистых растений Карпат показал, что жизнеспособность семян во многом определяется виталитет- ным состоянием особей, на которых они образуются.
С точки зрения зависимости уровня репродукции от состояния особей в популяции наиболее информативным является репродуктивное усилие. Репродуктивное усилие - это часть фитомассы растения, которую оно затрачивает на производство диаспор. Этот параметр обусловлен как состоянием особей, так и эколого -ценотической обстановкой и варьирует в достаточно широких пределах. Представляет интерес анализ зависимости репродуктивного усилия от фитомассы особей. У поликарпика частухи подорожниковойAlismaplantago-aquatica, поданным Н.В. Васильева (2006), репродуктивное усилие было меньшим у крупных особей. Мелкие особи тратили на репродукцию большую часть веществ, хотя фактически здесь реализовывалось простое соотношение: по ходу онтогенеза фитомасса особей прогрессивно нарастает, тогда как размер репродуктивных структур у генеративных растений состояний g{, g2 и g3 сравнительно стабилен.
Вычисляться репродуктивное усилие может разными способами: как фитомасса диаспор в расчете на единицу фитомассы растения или как фитомасса диаспор в расчете на единицу листовой поверхности. Как генетически обусловленный параметр репродуктивное усилие лучше вычислять в форме количества производимых диаспор в расчете на единицу фитомассы растений. Эта более по стоянная величина, отражающая прежде всего видовые особенное ти изучаемого растения (Репецкая, 2000). Репродуктивное усилие при его вычислении на единицу листовой поверхности особи, напротив, является более варьирующим показателем и к большей степени характеризует роль экосистемных факторов в репродукции. Как подчеркивал М.В. Марков (1990), определяться репрод у кг и с ное усилие должно в момент, когда репродуктивные и вегетативные органы имеют максимальное развитие, а в фитомассу репродуктивных органов следует включать и вспомогательные репродуктивные структуры (ось соцветия, прицветники и др.).
о
-5
50
%
40
ЗО
20
10
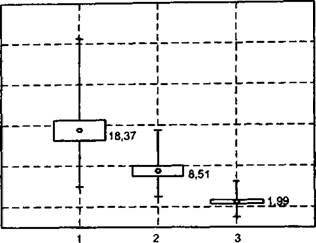
Рис. 2.7. Значения репродуктивного услилия(%) растений Polygonum scabrum в посевах разных сельскохозяйственных культур:
1 - узкорядный посев яровой пшеницы; 2 - посев озимой пшеницы; 3 - посев гречихи. Указаны значения среднего арифметического, значения среднего плюс/минус ошибка среднего арифметического (коробочки) и минимальное и максимальное значение величины репродуктивного усилия (усы)
На производство семян оказывают влияние не только со-т- л ние растений, ко и экологические условия на протяжении репродуктивного периода, конкурентные отношения с другими растениями сообщества, активность и численность вредителей и болезней растений (Fox et al., 1999). Для раскрытия роли этих факте ров могут использоваться различные статистическое мето о- приемы визуализации данных. Это иллюстрирует рис. 2. 7, на gt; , тором представлены значения репродуктивного усилия сорного растений горца шероховатого Polygonum scabrum в посевах трех разных сельскохозяйственных культур. При близкой средней численности сорняка (от 1,8 до 2,3 жт./м2) средние значені:а непродуктивного усилия в посевах разных культур были существенно различными, составляя в посеве яровой пшеницы - 18,37%, озимой пшеницы -8,51% и гречихи - 1,99%. Это свидетельствует о том, что подавление сорняка пологом этих культурных растений проявлялось не столько в снижении его численности, сколько в ингибировании репродукции.
Уровень популяций. На популяционном уровне репродукция растений характеризует количество производимых семян (диаспор), приходящееся на единицу поверхности популяционного поля. Этот параметр определяет репродуктивное давление популяции на экосистему.
Репродуктивная способность любой популяции зависит от двух факторов:
- репродуктивной производительности отдельных особей;
- количества особей на единицу площади, то есть плотности популяции.
Плотностная обусловленность репродукции состоит в том, что наибольшие репродуктивные выходы, как правило, связаны с некоторой средней плотностью популяции (Bgren, 1996). В маленьких популяциях нередко репродукция снижена из-за нехватки пыльцы, а в больших популяциях — из-за дефицита опылителей и ресурсов.
На уровне популяций полезно различать тычиночный и пестичный репродуктивный успех (Злобин, 1993). Первый состоит в производстве достаточного для опыления всех растений данной популяции количества полноценной пыльцы, а второй - в формировании полноценных семян в количестве, необходимом для поддержания стабильности популяции.
Ю.А. Злобиным (1968) была разработана категоризация урожая семян цветковых растений с выделением:
- потенциального урожая как количества семязачатков, закладываемых на генеративных растениях, в расчете ка единицу площади популяционного поля;
- фактического урожая как количества созревших и вступающих в процесс днссеминации семян или плодов;
- грунтового урожая как количества жизнеспособных семян в почве к моменту начала их возможного прорастания также з расчете на единицу площади.
Фактический урожай всегда оказывается ниже потенциального из-за недоопыления, абортирования семязачатков и их повреждения болезнями и вредителями, а грунтовый урожай меньше фактического из-за потерь при диссеминации, вызванных се- меядными животными.
Репродуктивное давление любой популяции на фитоценоз неравномерно по годам из-за чередования лет высокой и низкой семенной продуктивности. Такое чередование хорошо изучено у деревьев, проявляется оно и растений других жизненных форм (Herrera et ai., 1998). Периодичность семе ношений ведет к периодичности появления всходов, формируя «волны* и «взрывы» возобновления. Этот феномен оказывает большое влияние на итоговою возоэстную и онтогенетическую структуру популяций, ХОТЯ
і. практической работе многих исследователей он нередко упускается.
По числу производимых плодов и семян особи растений в популяциях неравноценны. Для осуществления репродуктивного процесса необходим некоторый критический минимальный раз меь особи (Yokoi, 1989). Размерные параметры матерински ч
особей особенно важны для участия в размножении у однолетних растений. Существует определенная взаимосвязь между размерами особей растений и их репродуктивным вкладом в самоподдер- жавие популяции. Как правило, крупные растения вносят в репродукцию существенно больший вклад. Но это нельзя рассматривать как универсальное правило. В популяциях нередко преобладают мелкие растения, и тогда их вклад в репродукцию оказывается сопоставимым с вкладом крупных растений, а то и превосходит его. В этой связи для репродуктивного потенциала популяций очень важна размерная структура особей, складывающих конкретную популяцию. Хорошо выражена зависимость репродуктивных показателей от возрастной, онтогенетической и вита- литетной структуры популяций. Репродукция выше в популяциях с высокой долей генеративных особей высшей категории вита- литета.
- Распространение семян и почвенные банки семян
Характер распространения семян и нераскрывающихся плодов во многом определяет структуру популяций и их динамику. С популяционной точки зрения особенно важно, что распространение семян имеет векторизированный характер и отличается огромным разнообразием форм и механизмов. Направление переноса у анемохоров определяется господствующими ветрами, у гидрохоров - направлением течения реки, у зоохоров - наличием и особенностями деятельности соответствующих групп животных и т.п. Во многих работах показано, что способы диссеминации приспособлены к условиям конкретного фитоценоза, к которому обычно приурочены популяции данного вида растения.
Важной особенностью растений является способность создавать запасы семян в почве, так называемые почвенные банки семян. Формирование запаса семян в почве происходит в течение длительного времени при обсеменении растений, входящих в состав фитоценозов, при заносе семян извне, за счет способности семян некоторых растений длительно сохранять всхожесть.
По реализации способности формировать почвенные банки семян все виды растений можно подразделить на три группа-
- семенной банк не создается, и семена прорастают в тот же вегетационный период, в котором они образовались;
- семенной банк носит переходный (сезонный) характер - семена зимуют и их основная масса прорастает весной следующего года;
- формируется истинный почвенный банк семян, сохраняющийся в почве в течение многих лет.
У рудералов почвенные банки жизнеспособных семян могут сохраняться десятки и даже сотни лет. Однако простой корреляции между способностью вида создавать почвенный банк семян и устойчивостью его популяций в фитоценозах не обнаружено (Sy mo nides, 1989).
В основе создания семенных банков лежит способность семян находиться в состоянии покоя. Обычно выделяется шесть типов покоя семян. В полевых исследованиях бывает достаточно трудно отличить покой семян, обусловленный генетическими, экологическими факторами или их взаимодействием. Очевидно только, что со временем размер почвенного банка семян (если он не пополняется) уменьшается по экспоненте (Movinchi etal., 2000).
Количество жизнеспособных семян в почвах, то есть размер почвенного банка семян, зависит от типа фитоценоза и характера почвы. По данным Т.В. Ивановой (2004), в почве луговых ценозов количество всхожих семян варьирует в пределах от 2027 до 7755 шт./м2, в поймах малых лесных рек размер почвенного бая ¦ ка семян меньше - 431-2232 шт./м2. В подмосковных лесах регистрируется до 750 шт. жизнеспособных семян березы на 1 кв. м. Е посевах количество семян сорняков в почве может достигать 400 ООО шт./м2, хотя обычно оно составляет 50 000-75 000 шт. Прослеживается тенденция к увеличению количества семян в пг*'' - венном банке от более сухих к более влажным сообществам. С нову почвенного банка семян во всех изученных сообществах чаще всего составляет небольшая группа видов-эксплерентов, на долю которых приходится более половины всей численности жизнеспособных семян в почве.
Формируются банки жизнеспособных спор И У ряда Я И? О а ровых растений. Но эта область популяционной экологии рас г; кий изучена мало.
Своеобразным экологическим резервом популяции являются и банки ювенильных и имматурных растений. Особенно они характерны для некоторых видов древесных пород (особенно дуб;., ели) когда подрост десятилетиями сохраняет жизнеспособное:* под пологом в виде «торчков*, а после возникновения окон в древостое ростовые процессы активизируются и торчки вырастают в полноценные деревья.
- Прорастание семян
Прорастание семян и выживаемость всходов составляют самостоятельную биолого-экологическую проблему, которой посвящено много публикаций, хотя большинство из них касается чисто фи- зиолого-биохимического аспекта (Fenner, 1985, Baskin, Baskin, 1998 я др. ). В популяционных исследованиях наиболее важно учитывать качество семян, то есть их способность прорастать и формировать полноценные всходы. Реализация этой способности зависит от многих условий. Большую роль играют температура, влага и для некоторых видов растений - свет. Разумеется, на прорастание семян влияют и другие факторы: аэрация субстрата, химические свойства почвенного раствора, деятельность животных, способных поедать семена и т.п. У некоторых видов растений прорастание стимулирует летнее прогревание семян, у других, напротив, зимнее промораживание. В этой связи было введено понятие безопасных микроместообитаний, или регенерационных ниш, где семена преимущественно прорастают, а проростки лучше сохраняются. Исключительно важным является срок прорастания семян, который часто играет критическую роль в выживаемости всходов.
У большинства растений максимум смертей приходится именно на фазу прорастающих семян. Уровень смертности прорастающих семян зависит от их качества и комплекса эколого-ценоти- ческих условий в период их существования и прорастания.
- Вегетативное разрастание и размножение
Как и в случае полового размножения, при анализе вегетативного размножения существуют два аспекта - уровень особей и уровень популяций.
Зачатки вегетативного размножения могут образовываться в разные периоды онтогенеза. Этот факт существенно влияет на их качество и жизнеспособность, так как от него зависит уровень
омоложения вегетативного потомства. Принято выделять три основных срока образования зачатков вегетативного размножения:
- Вегетативное размножение наблюдается в предгенеративный период или в самом начале генеративного размножения растения. В этом случае оно обычно сопровождается глубоким омоложением формирующихся партикул и будущих модулей. Ювенильная партикуляция приводит к образованию ювенильных или иммагурных дочерних особей (раметов) и играет особенно важную роль в самоподдержании популяций растений.
- Вегетативное размножение происходит в середине генератив ного периода и сопровождается лишь частичным омолаживанием. По Т.А. Рабогнову (1969), это нормальная партикуляция, которая ведет к образованию клоновых структур разного типа.
- Вегетативное размножение осуществляется в постгенератив- ный период. Это, по сути дела, сенильная партикуляция, или старческий распад, не связанные с глубоким омолаживанием потомства. Популяционная роль сенильной партикуляции невелика, она лишь продлевает срок контроля популяционного поля.
Конечно, такое подразделение вегетативного размножения носит самый общий характер, и у отдельных видов растений уровень омолаживания на том или ином этапе онтогенеза достаточно индивидуален. Так, у черники Vaccinium туrtUlus побеги расселения образуются в постгенеративный период, но уровень их омолаживания высокий, что ведет к формированию генераций жизнеспособных дочерних парциальных кустов и делает жизнь клона длительной (Злобин, 1962).
Для количественной характеристики вегетативного размноже яия, как и для генеративного, используются показатели потенциальная и реальная продуктивность диаспор вегетативного размножения.
В настоящее время особенности вегетативного размножения растений как популяционного феномена изучены еще очень слабо. У всех многолетних растений подземный корнепобеговый комплекс имеет очень большой резерв ювенильных меристем, от деятельности которых зависит восстановительная и регулирующая функция и особей, и популяций (Маслова и др., 2006). Его изучение - одна из актуальнейших проблем популяционной эколог ии.
Б.М. Миркиным и Т.Г. Горской (1989) было открыто важное яеяение. имеющее место е популяциях ращений, сочетающих
половое и вегетативное размножение. Оказалось, -что в этом случае между гаметами и раметами существует выраженная обратная связь. В формировании популяций таких растений при этом наблюдается три периода:
- инициальный, когда популяция и запас фитомассы в ней формируются за счет генетов. Это фаза первичного заселения территории;
- равновесный, когда в популяции устанавливается равновесие между генетами и раметами. В этот период доля генетов убывает, но это сразу же компенсируется нарастанием числа раметов;
- старения популяции, когда большая часть генетов из популяции выпали, а уровень вегетативного размножения затухает. В такой популяции преобладают стареющие раметы.
В целом, самоподдержааие в популяциях растений достигается разными способами и разными сочетаниями этих способов. Грайм (Grime, 1979) в этой связи выделил пяти типов репродуктивных стратегий:
- V-стратегия - вегетативное разрастание и размножение;
- S-стратегия - сезонное размножение семенами с весенним или осенним пиками их прорастания;
- Вг-стратегия - самоподдержание популяции за счет банка жизнеспособных семян в почве;
- Вр-стратегия - самоподдержание популяции за счет банка проростков и ювенильных растений, которые могут длительно сохраняться в фитоценозе;
- W-стратегия - возобновление за счет массово производимых семян или спор.
Именно многообразие форм и способов репродукции обеспе чивает популяциям растений стабильность и прогрессивное разрастание.
Еще по теме Самоподдержание и популяциях растений:
- Признаки популяций растений
- Популяции культурных растений
- Биоразнообразие особей растений в популяциях
- Сущность и формы линамики популяций растений
- Формы взаимоотношений между популяциями разных видов растений
- Эколого-ценотические стратегии вилов и популяций растений
- Разнообразие динамических процессов в популяциях растений разных жизненных форм
- Управление популяциями культурных и сорных растений
- Глава 7 Популяции споровых растений
- Глава 2 Особи растений - структурные единицы популяций
- ЗАКОНОМЕРНОСТИ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ. МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ ПОПУЛЯЦИЙ
- Границы между популяциями, иерархия популяций