РАДИОНУКЛИДНЫЙ СОСТАВ ЗАГРЯЗНЕНИЯ И ЗАКОНОМЕРНОСТИ НАКОПЛЕНИЯ Cs ТРАВЯНО-КУСГАРНИЧКОВЫМ ЯРУСОМ
По имеющимся немногочисленным данным, самоочищение травянистой растительности в среднем происходит в 3 раза интенсивнее, чем древесных пород [8]. Неадекватность снижения загрязненности древесного и травянистого ярусов связана с особенностями циклов развития растительности: однолетнего - для большинства трав и многолетнего - для деревьев. Произрастание травянистых растений и кустарников под пологом леса делает возможным и их вторичное загрязнение радиоактивными частицами, поступающими из древесного яруса в процессе его дезактивации (с чешуйками отшелушивающейся коры, продуктами вымывания осадков и т.д.). Все это как бы растягивает процесс самоочищения травяного покрова от поверхностного загрязнения и оказывает существенное влияние на динамику радионуклидного состава его загрязнения в целом.
По сравнению с ассимилирующими органами древесных пород в составе загрязнения травянистых видов более длительный период присутствуют такие радионуклиды, как 144Се, 95Zr, 95Nb и 106Ru, наличие которых в растениях в большей степени связано с внешним загрязнением (рис. 31).
Данные нуклиды в травянистой растительности регистрируются в течение 4-5 лет после выпадений. В дальнейшем в радионуклидном составе травянистых растений, как и ассимилирующих органов древесных пород, присутствуют только изотопы цезия и стронция.
Межвидовые вариации содержания радионуклидов в травянистых растениях более значимы по сравнению с таковыми у древесных пород. Это обусловлено следующими причинами: различиями в площади питания видов (у травянистых растений, в отличие от древесных, площадь питания невелика и не выходит за пределы отдельных форм микрорельефа); видовым разнообразием (у травянистых растений даже в пределах одного экотопа количество видов значительно превосходит видовой состав древесных пород).
Межвидовое варьирование содержания 137 Cs в травянистых растениях в среднем составляет около 45%, причем в течение вегетацион-
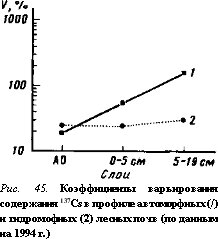
ного периода оно неодинаково и нарастает от весны к лету с последующим снижением к осени (табл. 38). Последнее, видимо, связано с сезонными вариациями накопления 137Cs в растениях.
Несмотря на очевидность связи между видовой принадлежностью травянистых растений и аккумуляцией в них радионуклидов, до сих пор не удается с достаточной убедительностью ранжировать отдельные виды по их накопительной способности [84, 85, 100, 175].
Возможность более достоверного ранжирования появляется при использовании величин средневзвешенных концентраций 137 Cs в фитомассе различных видов травянистого яруса за весь период вегетации.
На основании этого показателя среди доминирующих видов Украинского Полесья, развивающихся в пределах одного экотопа (ав-
Таблица 38. Межвидовое варьирование содержания 137Cs в некоторых видах травянистых растений уч. Д-1 (по данным 1989-1993 гг.), нКи/кг абс. сухой массы
|
Год |
Месяц |
п |
М |
±т |
G |
V, % |
max |
min |
|
1989 |
10 |
1,39 |
0,2 |
0,63 |
45,8 |
2,41 |
0,48 |
|
|
1993 |
Май |
7 |
0,33 |
0,05 |
0,15 |
46,4 |
0,52 |
0,09 |
|
Июнь |
7 |
0,28 |
0,06 |
0,17 |
61,5 |
0,67 |
0,11 |
|
|
Июль |
7 |
0,35 |
0,1 |
0,28 |
79 |
1 |
0,08 |
|
|
Август |
7 |
0,43 |
0,07 |
0,19 |
43,4 |
0,67 |
0,1 |
|
|
Сентябрь |
7 |
0,56 |
0,12 |
0,33 |
58,5 |
1,25 |
0,15 |
|
|
Октябрь 7 0,34 0,06 0,16 Примечание. У слов, обозн. см. табл. 26. |
47,9 |
0,56 |
0,11 |
|||||

томорфный ландшафт), выделены виды- концентраторы и дискриминаторы [82] или деконцентраторы по А.И. Перельману [179]. Среди растений-концентраторов по накопительной способности 137Cs выделяется ландыш майский (Convallaria majalis) и орляк (Pteridium aquili- num) (рис. 32).
Содержание 137 Cs в этих видах в 2-10 раз выше, чем в других растениях.
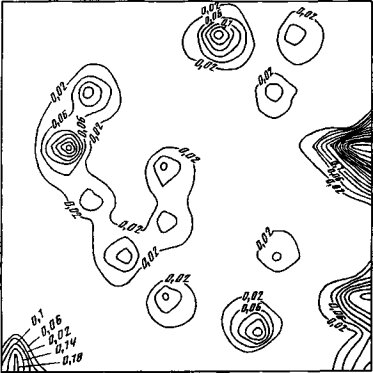
Значимое влияние на накопление радионуклидов травянистыми растениями оказывают условия их произрастания. В многолетних исследованиях прослеживается четкая закономерность увеличения содержания радионуклидов в растениях, произрастающих в условиях гидроморфизма (рис. 33). Кратность различий по этому признаку достигает 20-30 величин. На факторах, обусловливающих повышенное накопление радионуклидов растениями в гидроморфных условиях, уже неоднократно останавливались. Особое положение занимают растения-гигрофиты, в частности такие виды, как ирис водяной (Iris pseudacorus) и представители семейства осоковых (Сурегасеае). При одной и той же плотности загрязнения эти растения накапливают в сотни раз больше радионуклидов, чем виды-концентраторы на автоморфных почвах (см. рис. 33).
В структуре напочвенного растительного покрова максимальными уровнями накопления радионуклидов, в том числе 137Cs, характеризуются мхи (табл. 39).
Мхи, а также лишайники (в отличие от высших растений), обладают рядом свойств, приводящих к значительной аккумуляции в них химических элементов. Они довольно быстро реагируют на изменение концентрации радионуклидов в почве. Все это позволяет отнести мхи и лишайники к растениям-индикаторам радиоактивного загрязнения среды [27, 121, 162, 301, 309]. Накопление 137Cs мхами более, чем на порядок, выше величины его максимального накопления травянистыми видами и в 2-3 больше по сравнению с лишайниками. Причем, как правило, у мхов основная часть активности сконцентрирована в их отмершей части. Содержание 137Cs у плауновых почти такое же, что у папоротников, а наименьшее накопление радионуклидов в напочвенном
Таблица 39. Содержание 137Cs в мохово-лишайниковом и травянистом покровах в пределах одного эко топа, уч. Ш-5 (средние значения по данным на 1991 г.), кБк/кг
|
Вид |
137CS |
Вид |
137 CS |
|
Моховой покров |
Лишайниковый покров |
||
|
Dicranum rugozum: |
Cladina arbuscula |
407 |
|
|
вегетирующая часть |
703 |
Травяной покров |
|
|
отмершая часть |
1073 |
||
|
Pleurosiun schreberi |
Плауновые |
||
|
вегетирующая часть |
425,5 |
Lycopodium clavatum |
344,1 |
|
отмершая часть |
740 |
Папоротники |
|
|
Polytrichum |
Среднее |
332 |
|
|
juniperinum |
Цветковые растения |
||
|
вегетирующая часть |
703 |
Среднее |
146,2 |
растительном покрове характерно для высших цветковых растений. Отсюда очевидно, что мохово-лишайниковый покров является выраженным биогеохимическим барьером на пути вертикальной и горизонтальной миграции радионуклидов в лесных БГЦ.
Таким образом, использование показателей средневзвешенных концентраций радионуклидов в различных видах напочвенного покрова за весь вегетационный период позволяет дать более объективную оценку накопительной способности каждого вида по отношению к радионуклидам. Ранжирование растений, особенно хозяйственно-полезных, по их накопительной способности имеет большое практическое значение. Группа дискриминаторов может представлять интерес при подборе видов, пригодных для введения в культуру в зоне радиационного загрязнения, в то же время растения из группы концетраторов перспективны для изучения механизмов поглощения радионуклидов, а также могут быть использованы как биоиндикаторы при радиационном контроле сырья, заготовляемого на загрязненных территориях.
Еще по теме РАДИОНУКЛИДНЫЙ СОСТАВ ЗАГРЯЗНЕНИЯ И ЗАКОНОМЕРНОСТИ НАКОПЛЕНИЯ Cs ТРАВЯНО-КУСГАРНИЧКОВЫМ ЯРУСОМ:
- ОСОБЕННОСТИ ФОРМИРОВАНИЯ БЕРЕЗОВЫХ СООБЩЕСТВНА МЕЗОТРОФНЫХ ТРАВЯНО-СФАГНОВЫХ БОЛОТАХПОД ВЛИЯНИЕМ ОСУШЕНИЯ
- НАКОПЛЕНИЕ И ХРАНЕНИЕ НАВОЗА
- МЕСТА НАКОПЛЕНИЯ НАВОЗА
- НАКОПЛЕНИЕ ХОРОШО- И СИЛЬНОРАЗЛОЖИВШЕГОСЯВЕРХОВОГО ТОРФА В ЗАВИСИМОСТИ ОТ ВРЕМЕНИ НАЧАЛАБОЛОТООБРАЗОВАНИЯ
- Опыты по гибридизации растений. Накопление сведениЗ о наследуемых признаках
- Биологическая индикация загрязнения почвенной среды и самоочищения почв
- 4.6. ОХРАНА ВОДОИСТОЧНИКОВ ОТ ЗАГРЯЗНЕНИЯ
- Экологический контроль и рекультивация почв, загрязненных нефтью и нефтепродуктами
- 2.1. Донаучный период накопления знаний. Представления о «разуме» и «инстинкте» животных в трудах естествоиспытателей XVIII - первой половины XIX века
- ГИГИЕНА ПОЧВЫ И ОХРАНА ЕЕ ОТ ЗАГРЯЗНЕНИЯ
- СОДЕРЖАНИЕ ТЯЖЕЛЫХ МЕТАЛЛОВ И СЕРЫВ ФОНОВОМ И ТЕХНОГЕННО ЗАГРЯЗНЕННОМ БОЛОТАХ[3] Л. В. Карпенко
- ПРИЧИНЫ ЗАГРЯЗНЕНИЯ ПРИРОДНОЙ СРЕДЫ УДОБРЕНИЯМИ И ВОЗМОЖНЫЕ НЕГАТИВНЫЕ ПОСЛЕДСТВИЯ
- Способы ведения животноводства и принципы ветеринарной защиты в зонах загрязнения
- РАСТЕНИЯ-ИНДИКАТОРЫ ЗАГРЯЗНЕННОСТИ ОКРУЖАЮЩЕЙ СРЕДЫ
- Глава 10 ЭКОЛОГИЧЕСКАЯ ОЦЕНКА АГРОХИМИЧЕСКИХ СРЕДСТВ. ПУТИ ВОЗМОЖНОГО ЗАГРЯЗНЕНИЯ ОКРУЖАЮЩЕЙ СРЕДЫ УДОБРЕНИЯМИ
- Общие закономерности вертикальной стратификации микробных сообществ
- Общие закономерности выделения (экскреции) токсикантов из организма
- Геохимия -