Индукция диапаузы внешними факторами
Наиболее распространенный случай - факультативная диапауза, возникающая перед наступлением неблагоприятного сезона. Лишь у некоторых моновольтинных видов диапауза становится облигатной и ее наступление не зависит от условий.
Вполне логично было бы предположить, что основным фактором, контролирующим наступление диапаузы, является температура. Действительно, каждый сезон характеризуется своим уровнем температуры, а экспериментальные данные показывают, что есть определенная связь между уровнем температуры и наступлением диапаузы. Заметим, однако, что из года в год сезонный ход температуры несколько различен, а за теплой погодой может последовать резкое похолодание.
Другой возможный фактор, индуцирующий диапаузу, - это качество пищи. Так, мальвовая моль Gelechia malvella Hb. при питании молодыми завязями, цветами и листьями развивается без диапаузы, если же употребляет в пищу зрелые семена растений - всегда диапаузирует. Однако, диапауза имеет место в жизни очень многих насекомых, в том числе и тех, которые не питаются растительной пищей.
В 1920 г. американцы - химик В.Гарнер и физиолог растений Х. Аллард - опубликовали статью, ставшую важнейшим этапом в истории биологии. Они впервые показали, что организм способен реагировать на длину светового дня, изменяя в зависимости от нее характер своего развития. Перед исследователями была поставлена задача, которую мы назвали бы сейчас узко прикладной: почему один перспективный сорт табака, независимо от срока его посева, на севере США зацветает настолько поздно, что наступающие морозы убивают его семена. Исследователи перебрали все возможные варианты экспериментов, среди которых, конечно, температура стояла на первом месте, а длина светового дня - на одном из последних. Однако, оказалось, что если эти растения, выращиваемые при длинном летнем световом дне севера США, затенять, начиная с-16 ч дня, то они рано цветут и дают семена в летнее время, даже если стоит холодная погода. Гарнер и Аллард провели подобные эксперименты и с другими растениями и четко показали, что этапы развития растения определяются длиной светового дня.
Спустя три года вышла работа (S.Marcovitch, 1923), показавшая, что формообразование у тлей также определяется фотопериодом. Позже было обнаружено (M.Kogure, 1933), что эмбриональная диапауза у тутового шелкопряда определяется длиной светового дня, при которой развиваются яйца родительского поколения. В 1941 г. доцент кафедры энтомологии Московского университета Н. С. Андрианова нашла, что куколочная диапауза у китайского дубового шелкопряда Antheraea pernyi Guer. диктуется длиной светового дня, при которой выращиваются гусеницы.
И только, начиная с 50-х гг. нашего века, стало лавинообразно увеличиваться количество работ, описывающих фотопериодическую реакцию самых разнообразных насекомых. Здесь немалую роль сыграла школа, созданная заведующим кафедрой Ленинградского университета А. С. Данилевским (1961). В принципе, такого влияния светового дня на насекомых и следовало ожидать, поскольку длина светового дня зависит от астрономических причин и является самым точным и надежным сигналом, показывающим наступление того или иного сезона.
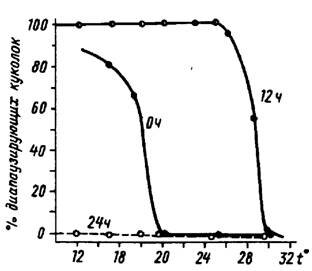
Рис.8. Зависимость доли куколок белянки Pieris brassicae L. в состоянии диапаузы от температуры и длины светового дня (по А. С. Данилевскому, 1961): 24 ч - постоянное освещение, 12 ч - короткий фотопериод, 0 ч - постоянная темнота
Влияние же других факторов обычно накладывается на фотопериодическую реакцию и ее модифицирует. Так, на рис.8 показана зависимость доли
диапаузирующих куколок бабочки капустницы от температуры при разной длине светового дня.
Таким образом, температура - важный фактор в индукции диапаузы, проявляющийся на фоне фотопериодизма. Возможны и случаи, как, например, у наездника-яйцееда Trichogramma pintoi Voegele., когда реакция на температуру более выражена, чем на фотопериод. Особенно важную роль играет температура в становлении диапаузы у таких объектов, которые обитают глубоко в почве или в древесине, куда практически не проникает свет.
Питание также существенно сказывается на становлении диапаузы, модифицируя влияние фотопериода. Так, у златоглазки Chrysopa cornea St. короткий световой день индуцирует диапаузу. Однако даже и при длинном световом дне недостаток тлей, которыми они питаются, приводит к проявлению диапаузы у 60-80% особей (MJ.Tauber et al., 1984). Аналогичным образом недостаток тлей в летнее время индуцирует диапаузу (эстивацию) у божьих коровок. Перед наступлением зстивации, как и перед зимней диапаузой, резко возрастает стремление этих жуков к миграциям (В.П.Семьянов, 1986). Как отмечалось выше, у фитофагов диапауза очень часто связана с изменениями биохимического состава растений в течение сезона (рис.9). В целом изменение пищи или ее недостаток чаще влияют на субтропических и тропических насекомых, где сезонные изменения светового дня не столь значительны.
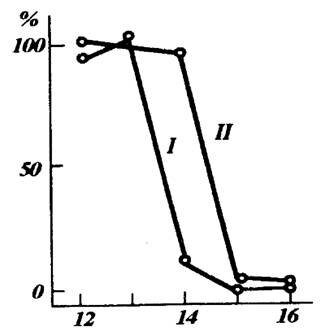
Рис.9. Фотопериодическая реакция хлопковой совки Helicoverpa armigera (Hubner) при питании гусениц разным кормом: / - коробочками хлопчатника, // - листьями хлопчатника. Ось абсцисс - фотопериод в ч, ось ординат - доля куколок в состоянии диапаузы (по Н.И.Горышину, 1958)
Большую роль в регуляции диапаузы может играть содержание влаги в пище и в окружающем субстрате. Так, диапауза яиц многих саранчовых продолжается до тех пор, пока не выпадет дождь. Подобная регуляция синхронизует выход личинок с появлением свежей растительности. Для начала эмбриогенеза у комаров Aedes также необходимо увлажнение. Известно, что самки этих комаров откладывают яйца по краям пересыхающих луж. Иногда наличием влаги определяется не диапауза, которая индуцируется фотопериодом, а возможность развития после диапаузы, как имеет место у лугового мотылька Loxostege sticticalis L.
Еще один важный фактор, способный сам по себе вызвать диапаузу, - это плотность популяции. Обычно высокая плотность способствует появлению диапаузы. Так, у насекомых - вредителей запасов (Ephestia cautella Wlk., Plodia interpunctella Hb.) скопление личинок, обычно возникающее при недостатке питания, приводит к их диапаузе, а в результате - к длительному сохранению популяции (H.Tsuji, 1963). Интересно, что у жуков-кожеедов Trogoderma variabile Ball, имеет место противоположный эффект. Личинки этого вида могут развиваться только в группах, у одиночных же личинок возникает диапауза (A.Elbert, 1979). Эффекты, связанные с плотностью популяции, а также с влажностью, далеко не всегда совпадают с определенным сезоном. Следовательно, диапауза - приспособление к любым неблагоприятным условиям, а не только к их сезонной смене.
Сигналы, индуцирующие диапаузу, могут быть различными даже у разных географических популяций одного и того же вида. Так, хлопковая моль Pectinophora gossypiella Saund., живущая в северной части своего ареала, реагирует на фотопериод, а в более южных районах - на температуру и качество пищи. Однако слабая реакция на фотопериод сохраняется и у популяций, обитающих около экватора (A.K.Raina and oth., 1981).
Еще по теме Индукция диапаузы внешними факторами:
- Внешние факторы
- Биотоп: местообитание и факторы внешней среды Местообитание и местопроизрастание
- РЕПРОДУКТИВНАЯ ДИАПАУЗА
- Индукция ферментов.
- 8.2.6. Эмбриональная индукция
- ДЕЗИНВАЗИЯ ОБЪЕКТОВ ВНЕШНЕЙ СРЕДЫ
- Роль внешней среды в формировании адаптаций
- Внешний осмотр тела птицы
- ГЛАВА 1. ВНЕШНЕЕ СТРОЕНИЕ
- 2.5. ТЕПЛООБМЕН МЕЖДУ ОРГАНИЗМОМ И ВНЕШНЕЙ СРЕДОЙ 2.5.1. ТЕПЛОПРОДУКЦИЯ Й ТЕПЛОПОТЕРИ ОТ ТЕШН№АТ5ЛРЪ?ЛРЕДЫ
- ВНЕШНЕЕ И ВНУТРЕННЕЕ СТРОЕНИЕ МЛЕКОПИТАЮЩИХ
- ВЛИЯНИЕ ВНЕШНЕЙ СРЕДЫ НА УСЛОВИЯ ПИТАНИЯ РАСТЕНИЙ И ЭФФЕКТИВНОСТЬ УДОБРЕНИЙ
- ВЛИЯНИЕ ВНЕШНИХ УСЛОВИЙ НА СПЕРМИЕВ ВНЕ ОРГАНИЗМА ЖИВОТНОГО
- ВЛИЯНИЕ ВНЕШНИХ УСЛОВИЙ НА СПЕРМИЕВ ВНЕ ОРГАНИЗМА ЖИВОТНОГО
- Главнейшие особенности внешнего строения насекомых других групп
- IV.2. ПОГЛОЩЕНИЕ АЭРОИОНОВ ВНЕШНЕГО ВОЗДУХА РАЗНЫМИ ФИЛЬТРАМИ И ПРИ КОНДИЦИОНИРОВАНИИ
- РЕМАТОДОЗЫ Систематика, морфология и биология трематод. Морфофункциональная характеристика внешних покровов (тегумента) трематод
- НЕМАТОДОЗЫ Систематика, морфология и биология нематод. Морфофункциональная характеристика внешнего строения нематод