Шельф
Как указывает A. Longhurst (1998) донная фауна Гвинейского залива на поверхности 28°, а на глубине, не превышающей высоту деревьев тропического леса всего 16°С. При этом видовой состав биоты края шельфа Гвинейского залива весьма сходен с таковым Западной Европы, а температура отличается не более, чем на 2°С, тогда как видовой состав и абиотические условия верхней сублиторали Гвинейского залива и Западной Европы различаются весьма существенно.
А.Н. Голиков указывает, что «северная граница Японской низкобореальной подобласти проходит в районе м. Терпения и северной части о. Итуруп. Однако глубины свыше 30 м в заливах Анива и Терпения занимает Алеутская высокобореальная подобласть» (1982: 97). Поэтому не удивительно, что рассмотрение всего шельфа как единого биогеографического региона весьма существенно искажает реальную картину. Возможно, пытаясь отразить неоднородность биот по глубине, но оставаясь в рамках «биогеографии всего шельфа» А.Н. Голиков (1980, 1982) вводит в биогео- графическую характеристику отдельных видов данные по глубине их обитания, что при выделении действительно бмогеографических регионов было бы совершенно излишне.Тем не менее, несмотря на то, что многочисленные данные свидетельствуют о существовании на шельфе, по крайней мере в умеренных и тропических водах, двух сменяющих друг друга по вертикали биот, районирование проводили и проводят для всего шельфа. В первом приближении для обзорных целей можно считать фиталь соответствующей верхней сублиторали. Поэтому схему районирования по макрофитам можно рассматривать как схему районирования верхней сублиторали. Более того, поскольку в потоках энергии верхней сублиторали существенную роль играет фотосинтез донных макрофитов и рифов герматипных кораллов, эти организмы являются эдификаторами соответствующих ассоциаций, часть из которых являются климаксными. Поэтому схемы районирования по макрофитам и герматипным кораллам должны хорошо соответствовать схеме районирования верхней сублиторали в целом, по крайней мере сообществ твёрдых грунтов.
В первую очередь для карты районирования по макрофитам (рис. 6.6) специфично разделение всей верхней сублиторали на 5 областей: 2 тропических (индо-тихоокеанскую и атлантическую), 2 умеренных (бореальную и аустральную) и антарктическую.
Разделение тропического пояса на два региона подтверждают данные по другой группе-эдификатору верхней сублиторали — герматипным кораллам (см. Главу 3, раздел «Коралловые рифы»).
Как показано в упомянутом разделе, граница между ЦС, эдификаторами которых являются макрофиты и рифы герматипных кораллов имеет биотическую природу: в более высоких широтах на твёрдых субстратах более конкурентоспособными являются ЦС с климаксом, образованном макрофитами, преимущественно бурыми водорослями, а в низ-
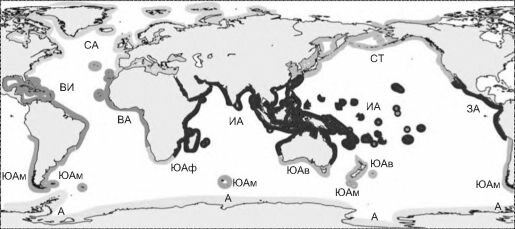
Рис. 6.6. Фитогеографическое районирование Мирового океана по макрофитам Ю.Е. Петрова (1974).
Одной заливкой дана одна область, подобласти обозначены буквами. Бореальная область: СА — Североатлантическая подобласть; СТ — Северотихоокеанская подобласть; Тропико-атлантическая область: ВИ — Вес- тиндская подобласть; ВА — Восточноатлантическая подобласть; Индо-пацифическая тропическая область: ИА — Индоавстралийская подобласть; ЗА — Западноамериканская подобласть; Аустральная область: ЮАм — Южноамериканская подобласть; ЮАф — Южноафриканская подобласть; ЮАв — Южноавстралийская подобласть; Антарктическая область — А.
ких — герматипными кораллами, а на литорали — манграми. При сравнении рис. 6.6 и рис. 3.56 бросается в глаза также совпадение границы тропической зоны с границей распространения рифов герматипных кораллов. Это позволяет сделать вывод, что в верхней сублиторали это взаимодействие определяет положение границы между тропическими областями и субтропическими регионами. Иными словами, на границе тропиков происходит замена доминирующего таксона климаксных ассоциаций и даже способа фотосинтеза: герматипные кораллы — в тропиках, крупные бурые макрофиты-водоросли — в субтропиках, умеренных и холодных водах.
О причинах этого в настоящее время мало что известно. Необходимы исследования как самого симбиоза, так и сравнение биохимии и энергетики фотосинтеза симбиотрофных организмов и макроводорослей, а также биотических взаимоотношений в экосистемах верхней сублиторали. На современном уровне знаний можно отметить следующие моменты:- В тропиках симбиоавтотрофами являются не только герматипные кораллы и другие Cnidaria, но и моллюски и губки, реже — другие группы. При этом с увеличением широты количество симбиотрофных видов и средняя степень развития симбиотрофии падает. Рассмотрим в качестве примера уже упоминавшихся ранее при анализе Сарматской биоты двустворчатых моллюсков Cardiidae. Это семейство, насчитывающее несколько сотен видов, обладает космополитическими распространением на шельфе, хотя в высоких широтах число видов и невелико (на арктическом шельфе — 2), всё же целое подсемейство Clinocardiinae обитает только в умеренных и холодных водах Северного полушария. Буль- шая часть видов питается, фильтруя придонный слой воды и детрит, лежащий на поверхности грунта. 9 видов специализированных Tridacninae (иногда рассматриваемых как самостоятельное семейство) обитают только в тропиках. Среди них крупнейшие современные двустворчатые моллюски, достигающие размера 137 см и веса более 500 кг (Tridcicnci gigas). Мантия тридакн содержит большое число симбиотических водорослей. Сама тридакна лежит на дне на коралловых рифах макушкой вниз, развернув и выпятив мантию так, чтобы она занимала максимальную поверхность. 95% синтезированного органического вещества симбиотические водоросли передают хозяину (Flitt, 1993). Продукции симбиотических водорослей, по-видимому, хватает для покрытия по крайней мере большей части энергетических потребностей тридакн, т. е. они занимают нишу фотосинтетиков, хотя все тридакны способны и фильтровать. Это позволяет тридакнам расти со сказочной быстротой — до 10 см в длину в год, при этом толщина створок увеличивается на 1 см в год! Между этими двумя крайними вариантами питания среди кардид (детритофагия и симбиоавтотрофия) имеются и виды со смешанным питанием. Даже разные виды Tridcicna удовлетворяют свои потребности за счёт симбиотических водорослей в разной степени. Виды, имеющие симбиотических водорослей, дающих «прибавку» к рациону, но не удовлетворяющих и половины потребностей, есть среди Fragiinae и даже Clinocardiinae, причём симбиоз этот не столь облигатен. Имеется тенденция снижения числа видов с развитым в той или иной степени симбиозом с водорослями по направлению к полюсам. По-видимому, фототрофное питание с помощью симбиотических водорослей выгодно для вида (энергетически?) только в низких широтах.
- Крупные макрофиты требуют для своего функционирования непрерывного приноса биогенов и уноса продуктов метаболизма. Эти проблемы вовсе не стоят перед герматипными кораллами, поскольку, как отмечалось выше, они способны фотосинтезировать при содержании фосфора в окружающей их воде на уровне лимитирования фотосинтеза фитопланктона, а азот симбиотической водоросли поставляют кораллы, усваивающие и продукты обмена водоросли. Заросли крупных бурых макрофитов (саргассы, Turbinaria) в тропиках развиваются только при разрушении коралловых рифов, как первая стадия сукцессии. Через несколько лет кораллы вытесняют водоросли. Ламинариевые в тропиках отсутствуют полностью.
- В тропическом поясе обитает много видов, питающихся макрофитами, с продвижением в высокие широты число таких видов резко и быстро падает. Например, рыбы и морские ежи, специализирующиеся на питании морскими травами, отсутствуют в боре- альных водах, хотя многочисленны в тропиках (Vermeij, 1978). Устранение видов, питающихся макрофитами, в эвтрофных районах приводит к бурному развитию водорослей и отмиранию кораллов. Такая смена происходит в прибрежных водах Вьетнама, где местное население выловило практически всех рыб и крупных беспозвоночных для продажи и питания (личн. сообщ. Д. Астахова).
Что касается зообентоса, то районирование здесь проводят в целом для шельфа, поэтому дать отдельно схемы районирования верхней и нижней сублиторали не представляется в данный момент возможным.
Часто биогеографические схемы пытаются совместить со схемами широтной зональности. Выделяемые зоны имеют характер поясов и при их выделении широко используют гидрологические критерии. Поэтому, строго говоря, эти зоны в действительности являются, в противоположность тому, что утверждают их авторы, не биогеографическими, а климатическими, отражающими широтную зональность в распределении поступления солнечной энергии на Землю, что приводит к широтной зональности в распределении температуры, освещённости и др. Конечно, распределение жизни на Земле определяют и абиотические факторы, в частности температура, но сущность схемы определяет логическое основание: распределение абиотических факторов (например, температуры) или распределение жизни.
«Разные авторы выделяют между Северным полюсом и экватором три (Зенкевич, 1948; Зёрнов, 1949), четыре (Hedgpetli, 1957; Peres, 1961; Briggs, 1974), пять (Coomans, 1962), шесть (Stephenson, 1947; Hall, 1964; Кусакин и др., 1975), семь (Зинова, 1962), девять (Семёнов, 1977) зон и подзон» (Несис, 1985: 54).
Авторы схем широтной зональности пытаются найти закономерности в смене биот от экватора к полюсам путём выделения широтных зон, имеющих глобальное значение. Эта идея основана на наблюдаемом в масштабе планеты градиенте «экватор - полюса». Эту ось и пытаются разбить на отрезки, соответствующие наиболее крупномасштабным биогеографическим выделам. Это легко сделать вдоль одного берега океана, но подобная схема фактически является повторением обычной частной биогеографической схемы и в её существовании немного глобального смысла. Сложности начинаются при попытке свести схемы, построенные от каждого полюса к экватору вдоль каждого берега материка, в одну общую. В разных схемах широтной зональности общими являются только эти две опорные точки: экватор и полюс.
«Произвольное использование в качестве объекта изучения биогеографии наряду с ареалом условий обитания в одних случаях исказило выявленные биогеографические закономерности, в других — просто помешало их выявить.
В настоящее время нет ни одной системы, отражающей зональную дифференциацию морских организмов, которая в большей или меньшей мере не грешила бы этим недостатком» (Перестенко, 1982: 100).«Физико-географическая, ландшафтная «заданность», предопределённость вопреки свойствам самой фауны очень характерна для сторонников ландшафтно-зональной концепции фаунистического комплекса» (Чернов, 1984: 739).
Количество же широтных зон вдоль каждого берега каждый биогеограф волен выбирать в зависимости от его представлений и методов, что, собственно говоря, и делают. Ещё больше возможностей представляется при проведении широтной зональности в пределах отдельного океана и ещё больше — в пределах всего Мирового океана. Поскольку нет общепринятых методов ни районирования, ни сравнения выделенных районов и их группировки, в литературе наблюдается достаточно большое разнообразие схем широтной зональности, особенно, если учесть малое число исследователей, занимающихся их построением. При этом обычная практика состоит в объединении регионов, расположенных в сходных условиях, а не населённых сходными биотами, проведение границ между зонами в местах наиболее резких градиентов абиоты, а не смены биот. До некоторой степени это обусловлено тем, что, как отмечал К.Н. Несис (1982: 115), «сопоставление схем для противоположных сторон океана и разных океанов будет фаунистически обосновано лишь при значительном числе общих — амфиокеанических — видов в контралатеральных провинциях. Число таких видов велико в приполярных областях, где преобладают циркумглобальные ареалы, и прогрессивно снижается при движении к экватору». Положение осложняет то, что протяжённость ареала одного и того же таксона вдоль разных берегов разных океанов обычно различна (см., например карту ареала морских ежей рода Brisaster, рис. 3.7, краба Chionoecetes opilio, рис. 4.43 и полихеты Nothria hyperborea, рис. 5.9), поэтому в разных районах он входит в состав разных широтных поясов.
Лишь немногие авторы честно пишут, что проводят широтную зональность, ориентируясь на абиотические факторы: «широтная зона — результат объединения районов, выделенных по фаунистическим показателям, но объединённых по абиотическому признаку» (Миронов, 1981: 12). Как абиотический признак часто используют температуру, например среднемесячные поверхностные изотермы самого тёплого месяца 25°С, 20°С, 15°С и 10°С для проведения границ, соответственно, тропического, субтропического, тёплого умеренного, холодного умеренного и арктического поясов. Очевидно, хорошего совпадения этих изотерм с биогеографическими границами ожидать не следует, и действительно, авторам приходится объяснять эти отклонения или использовать в каждом случае (например, для объяснения положения границ у разных берегов одного океана) разные показатели.
При проведении действительно биологического районирования Мирового океана следует учесть, что в каждом из минимальных биогеографических выделов (районов) есть как минимум две сукцессионные системы со своими климаксами: сукцессионная система рыхлых грунтов и сукцессионная система твёрдых грунтов. Обоснование этого было дано в главе «Синэкология», здесь же целесообразно остановиться на следствиях.
Прежде всего, надо отметить, что к ассоциациям твёрдых грунтов относится как бульшая часть ассоциаций макрофитов, так и все мелководные коралловые рифы, т. е. рис. 6.6, как и рис. 3.56 — это схемы биогеографического районирования преимущественно ассоциаций твёрдых грунтов.
Рыхлые грунты с точки зрения биоты гораздо менее гетерогенны, нежели твёрдые (скалы, валуны). На них виды могут расходиться, прежде всего, по типу питания (фильтрация придонного слоя воды на разной высоте от грунта, питание поверхностным или подповерхностным детритом, хищничество) и степени углубления в грунт. Часто отмечаемая связь распределения отдельных видов с преобладающим размером фракции грунта (гранулометрией), вполне возможно, является отражением сукцессионной динамики и/или локальных условий местности, в которой проводился анализ, т. е. обусловлена не гранулометрическим составом грунта самим по себе, а неучитываемыми в анализе локальными факторами, связанными с ним. При анализе данных, собранных в гораздо буль- шем районе, локальные факторы теряют свою значимость и результаты получаются совсем иными. Так, мы на материале бентосных съёмок Баренцева моря 1968-1970 гг. проанализировали связь распределения разных видов полихет-обитателей рыхлых грунтов одновременно с гранулометрическим составом грунта, глубиной и температурой в масштабах всего моря на основании проб с 639 станций. Достоверная связь распределения с гранулометрическим составом грунта не обнаружена для многих видов, обитающих и питающихся в толще грунта. В то же время, анализ показал значимое взаимодействие абиотических факторов между собой, в частности глубины и гранулометрического состава грунта — на уровне 99,9%.
На твёрдых грунтах возможно расхождение также по неоднородностям субстрата (под камнями, в щелях, на поверхности камней, в норах, вытачиваемых в субстрате и т. д.). Поскольку характер этих условий резко меняется в пространстве, у обитателей твёрдых грунтов гораздо чаще встречаются разные формы территориального поведения. Бульшая гетерогенность биотопа приводит к тому, что сообщества твёрдых грунтов включают бульшее число видов, поэтому сравнение суммарного состава биоты биогеографического
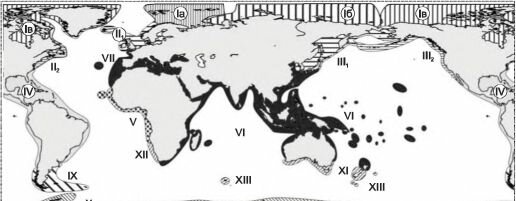

Рис. 6.7. Зоогеографическое районирование шельфа Е.Ф. Гурьяновой (1964).
I — арктическая область: 1а — баренцевоморская провинция; 16 — сибирская провинция; 1в — чукотско-американская провинция; II — бореальная северо-атлантическая область: Щ — западно-европейская подобласть; ІІ2 — северо-американская подобласть; III — бореальная северо-тихоокеанская область: IIIі —дальневосточная подобласть (беринговоморская провинция; охотоморская провинция; северо-западная япономорская провинция, юго-восточная япономорская провинция, курильская провинция); ІІІ2 — орегонская подобласть; IV — тропическая область Центральной и Южной Америки; V — западно-африканская, или гвинейская тропическая область; VI — индийско-западно-тихоокеанская тропическая область (Индо-вест-пацифичес- кая); VII — лузитано-средиземноморская фауна индийско-западно-тихоокеанской области; VIII — северо-чи- лийская область; IX — антибореальная область; X — антарктическая область; XI — южно-австралийская, или тасманская область; XII — южно-африканская область; XIII — эндемичные фауны субантарктических островов.
региона будет в большей степени отражать сходство/различие сукцессионных систем твёрдых грунтов. Виды, входящие в состав сообществ твёрдых грунтов, имеют меньший геологический возраст, нежели виды, входящие в состав сообществ рыхлых грунтов. G.J. Vermeij (1978) проанализировал скорость видообразования в мелководных сообществах по разные стороны Панамского перешейка и обнаружил, что число морфологически сильно разошедшихся видов существенно выше среди видов твёрдых грунтов, чем рыхлых. До образования перешейка этот район был населён единой биотой. Нет оснований полагать, что климатические изменения, происходившие в изучаемых районах, влияли по- разному на обитателей рыхлых и твёрдых грунтов. Поэтому различия в числе морфологически сходных или идентичных видов, обитающих по разные стороны перешейка на разных грунтах, т. е. степень морфологического расхождения исходного вида по разные стороны перешейка, можно считать функцией скорости эволюционных преобразований.
Различия в видовом богатстве сукцессионных систем рыхлых и твёрдых грунтов одного биогеографического региона и различия в скорости их эволюции, возможно, следует учесть в дальнейшем при анализе структуры биосферы.
На рис. 6.7-6.11 приведены несколько карт биогеографического районирования шельфа Мирового океана. Эти схемы, как и схему на рис. 6.5, для удобства сравнения я перерисовал, при этом один биогеографический выдел крупного ранга заштрихован одинаково, более крупного ранга — однотипно (например, область заштрихована, подобласти показаны разной штриховкой, другая область залита, подобласти показаны заливкой разной интенсивности и т. п.) и, по возможности, сходно на разных схемах. Более мелкие выделы обозначены буквенными или цифровыми индексами (как в оригинале).
Первая из схем (рис. 6.7) составлена Е.Ф. Гурьяновой, в ней она попыталась отразить генетические взаимоотношения фаун. Так, например, она объединила в один регион тропические районы восточной Пацифики и западной Атлантики (IV на рис. 6.7), а также отнесла восточно-атлантический регион от Ла-Манша до о-вов Зелёного мыса, включая
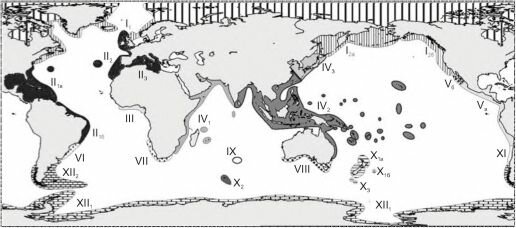
Рис. 6.8. Зоогеографическое районирование шельфа О.Н. Зезиной (1971).
- — бореально-арктическая область: 1\ — северо-атлантическая подобласть; І2 — северотихоокеанская подобласть; 12а — азиатско-алеутская провинция; I2g — североамериканская провинция; I3 — арктическая подобласть;
- — афмиатлантическая область; Их — атлантическо-центральноамериканская подобласть: II|a — карибская провинция; IIхб — бразильская провинция; П2 —лузитано-мавританская подобласть; II3 — средиземноморская подобласть; III —западноафриканская область; IV — индо-западнотихоокеанская область: IVх —западноиндоокеанская подобласть; IV2 — индомалайская подобласть; IV3 — японская подобласть; V — тихоокеанско-центральноамериканская область: Va — центрально-американская провинция; Уб — калифорнийская провинция; VI — южнобразильско-уругвайская область; VII — южноафриканская область; VIII — южно-австралийская область; IX — ново-амстердамская область; X — новозеландско-кергеленская область: Хх — новозеландская подобласть; Х1а — северо-новозеландская провинция, Ххб — южно-новозеландская провинция; Х2 — кергеленская подобласть; Х3 — маккуорианская подобласть. XI — чилийский переходный район; XII — антарктическо-южно- американская область: ХЩ — антарктическая подобласть; ХП2 — южно-американская подобласть.
Средиземное море (V на рис. 6.7), к Западно-Индо-Тихоокеанской области. И то, и другое никем признано не было, поскольку не подтверждается фактическими данными. Особенно это касается отнесения района Средиземного моря и прилежащих районов к Индо-западно-тихоокеанской области. В действительности от биоты Тетиса в Средиземном море ничего не осталось (оно с момента исчезновения Тетиса неоднократно высыхало). Реликтов биоты Тетиса, равно как и их потомков в Средиземном море и прилежащих районах меньше, чем в западной Атлантике. Относительно предлагаемого разделения шельфа Арктики на три региона — см. ниже. При построении иерархии регионов Е.Ф. Гурьянова использовала метод соответствия ранга регионов рангу эндемичных для них таксонов, который, как показано в предыдущей главе при характеристике этого метода, на практике использован быть не может.
Вторая схема (рис. 6.8) построена О.Н. Зезиной по брахиоподам. Регионы были выделены методом типизации ареалов и сгруппированы методом оценки сходства списков. К сожалению, в качестве показателя ранга был выбран индекс Престона, который, как показано выше, не следует использовать (хотя это и широко делают), в результате ранг выделенных районов фактически не обоснован.
Третья карта (рис. 6.9) скомпилирована из отдельных карт монографии J.C. Briggs (1995). В основу схем J.C. Briggs положил анализ распространения всех организмов бентоса и рыб. Карта J.C. Briggs’a отличается наиболее детальной проработкой, но обоснованность выделенных районов и их ранг часто вызывают сомнение, и, наоборот, в отдельных случаях степень отличия биот не находит адекватного отражения. Причина в том, что J.C. Briggs считал отдельной провинцией такую акваторию, которая имеет 10% эндемичных видов (Briggs, 1995: 231). Как показано в предыдущей главе, критерий доли эндемичных видов можно использовать только при составлении самых первых, прикидочных грубых схем. К тому же, он выделял акватории для сравнения, по-видимому, в значительной степени по гидрологическим данным. Это явствует, во-первых, из того, что наиболее
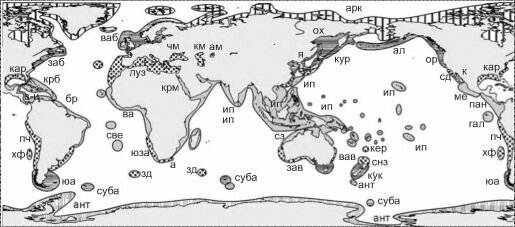
Рис. 6.9. Биогеографическое районирование шельфа J.C. Briggs (1995).
Тропические регионы: индо-вестпацифический (провинции: крм — Красного моря, зи —западного Индийского океана, ип — индо-полинезийская, сза — северозападная австралийская), восточный тихоокеанский (провинции: ме — мексиканская, гал — галапагосская, па — панамская), западный атлантический провинции: крб — ка- рибская, бр — бразильская, в-и — вест-индийская, све — островов Святой Елены), ва — восточный атлантический.
Субтропические (warm-temperate) регионы: средиземноморско-атлантический (провинции: луз — лузитан- ская, чм — Черного моря, км — Каспийского моря, ам — Аральского моря); южноавстралийский (зав — западноавстралийская и вав — восточноавстралийская провинции), североновозеландский (снз — североновозеландская и кер — кермадекская провинции); Западный южноамериканский (провинции: пч — перуано-чилийская и хф — островов Хуан-Фернандес); восточный южноамериканский; южноафриканский (провинции: юза — юго- западно африканская, а — Агулхаская (Agulhas), зд — западного дрифта); кар — каролинский; калифорнийский (провинции: сд — Сан-Диего и к — Кортеса); я — японский.
«Экман и Бриггс использовали в своих схемах зонально-географический принцип и данные о важнейших климатических границах в морской среде, которым соответствует их границы между областями («Regions»), и не заметили одну из самых существенных границ у берегов Южной Америки — в районе Огненной Земли и Фолклендских (Мальвинских) островов» (Семёнов, 1982: 215).
Умереннохолодноводные (соId-temperate) регионы: западнотихоокеанский (провинции: ор — ориентальная, ох — охотоморская и кур — курильская); юа — южно-американский, та — тасманский; южный новозеландский (провинции: кук — островов Кука и ант — островов Антиподов); восточнотихоокеанский бореальный (провинции: ал — алеутская и ор — орегонская); заб — западноатлантический бореальный; ваб — восточноатлантический бореальный; суба — субантарктический (несколько провинций на изолированных группах островов) Полярные (cold, или polar) регионы: арк — арктический и ант — антарктический.
крупные регионы его схемы соответствуют регионам, выделенным по температурному режиму, а. не по различиям в биоте. Так, он считает равными ранг арктического района и умеренных вод северных Атлантики и Пацифики. Отечественные авторы, более знакомые с этими регионами, или выделяют три равноценных региона для Арктики, умеренной Атлантики и умеренной Пацифики (Е.Ф. Гурьянова) или объединяют арктический регион с каким-либо умеренным (O.H. Зезина,
К.Н. Несис). Во-вторых, J.C. Briggs не проводил биогеографические границы в районах резкой смены биоты, если там нет резких гидрологических градиентов (юг Южной Америки, южная Норвегия). Поэтому схема J.C. Briggs’a в действительности не биогеографическая, а эклектичная биогеографо-климатическая.
Четвёртая схема (рис. 6.10) предложена К.Н. Несисом (1982а,б), преимущественно на основе анализа распространения головоногих моллюсков. Регионы выделены методом типизации ареалов, но самостоятельными признаны только регионы, имеющие эндемичные виды (не обязательно головоногих моллюсков). На этой схеме выделено всего три области: одна тропическая и расположенные по обе стороны от неё бореально-арктическая и нотально-антарктическая. Кроме того, выделена одна (две?) надпровинции, а границы
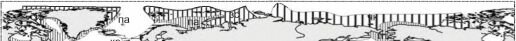
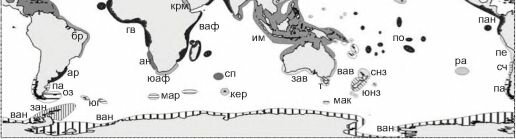
Рис. 6.10. Зоогеографическое районирование шельфа К.Н. Несиса (1982а, б). Тропическая область с подобластями: западноатлантическая (провинции: кар — каролинская, ви — вестинд- ская, бр — бразильская, ар — аргентинская), восточноатлантическая (гв — гвинейская, ан — анголо-намибская, юаф — южноафриканская), индо-вестпацифическая (крм — красноморская, ваф — восточноафриканская, им — индомалайская, юя — южнояпонская (сино-японская), г — гавайская, по — полинезийская, сп — сенпольская, зав — западноавстралийская, вав — восточноавстралийская, снз — североновозеландская, р — рапануйская), восточнотихоокеанская (кал — калифорнийская, пан — панамская, пе — перуанская).
Бореально-арктическая область с подобластями арктической (зс — западносибирская, чк — чукотско-канадская, па — приатлантическая переходная, пт — притихоокеанская переходная), атлантической бореальной (акд — акадийская, ке — кельтская (скандинавская), вир — виргинская), тихоокеанская бореальная (ак — алеутско-камчатская, ся — северояпонская (айнская), ор — орегонская).
Лузитано-средиземноморская надпровинция (л — лузитанская, ср — средиземноморская, мав — мавританская). Нотально-антарктическая область с подобластями: Магеллановой (сч — среднечилийская (арауканская) переходная, пат — патагонская, оз — огнеземельская), кергеленской (мар — марионская, кер — кергеленская, мак — маккуорийская), антарктической (юг — южногеоргийская, зан — западноантарктическая (антарктандийская), ван — восточноантарктическая (антарктгондванская).
Тасманийская (т) и южноновозеландская (юнз) провинции не включены ни в одну область, их положение не обсуждается, вероятно, они, аналогично лузитано-средиземноморской, образуют отдельную надпровинцию.
На оригинальной схеме области названы суперригионами и подобласти — регионами; провинциии обычно разделены более или менее широкими переходными зонами, на данной её копии переходные зоны оставлены без карто граф иче ско й нагрузки.
между регионами не резкие, а в виде переходных зон. И то, и другое, на мой взгляд, является артефактом использованного подхода к выделению регионов: изначально фаунисти- ческие различия верхней сублиторали и элиторали были признаны биогеографически незначимыми. В действительности, в пограничных районах (и в Средиземном море) более южные биоты обитают в верхней сублиторали, тогда как более северные — в элиторали. Совместное их рассмотрение и создаёт иллюзию смешанности биоты.
Последняя схема (рис. 6.11) представляет собой составленную мною компиляцию различных схем районирования шельфа. Компиляция была сделана на основе идеологии, описанной в двух предыдущих главах. Несколько замечаний к схеме:
- Схема доведена только до уровня провинций, хотя это и не самое мелкое подразделение. Выделение ареалов ЦС (биогеографических районов) на данной стадии развития биогеографии моря возможно только в очень хорошо изученных в этом отношении районах (см. ниже районирование Северного Ледовитого океана).
- В одних случаях различные использованные для компиляции схемы давали хорошее совпадение, в других они более или менее существенно различались, особенно в числе районов. При составлении своей компиляции я использовал наименее дробные схемы, поскольку дробность более детальных схем не всегда была на мой взгляд достаточно обоснована.
- В то же время, по-видимому, в большинстве провинций следует разделить шельф на верхнюю сублитораль и элитораль, удалось это сделать только в пределах бореальных районов Северного Ледовитого океана (на арктическом шельфе это разделение, видимо, отсутствует). В любом случае, реальная дробность, конечно, выше, чем приведённая на моей схеме. При этом совершенно не обязательно совпадение границ между районами верхней сублиторали или элиторали. Вероятно, здесь также, как и при переходе от шельфа к батиали и далее к абиссали, будет наблюдаться увеличение размеров биогеографических регионов. Особенно не удовлетворяет меня районирование наших Дальневосточных морей. Однако все предложенные ранее схемы, на мой взгляд, судя по имеющимся у меня данным по распространению полихет и учитывая распространение других групп (двустворчатых моллюсков, иглокожих и др.), также весьма далеки от реальности, в первую очередь из-за совместного рассмотрения всего населения шельфа и, нередко, даже и склона. На данный момент я не в состоянии предложить аргументированную схему районирования, поэтому весь этот район отнесён к одной провинции.
- Районирование Северного Ледовитого океана дано по моим предыдущим работам (Жирков, 2001, 2004).
- Районирование Южной Америки дано по В.Н. Семёнову (1982).
- В тропическом поясе, основываясь на данных по макрофитам и коралловым рифам, выделено две области: атлантическая и индо-пацифическая.
- Нет разделения бореальных районов у атлантического побережья Северной Америки, традиционно проводимого биогеографами школы ЗИН, здесь я следую последним данным американских авторов (Briggs, 1999), а также биогеографам московской школы.
- Каспийское море рассмотрено как регион ранга области (Зенкевич, 1947; Морду- хай-Болтовской, 1960). При этом солоноватоводной биотой, как показано выше, населена только солоноватоводная провинция, к которой отнесены Северный Каспий и распреснён- ные районы Черного и Азовского морей. Средний и Южный Каспий выделены в особый биогеографический регион, населённый морской биотой, сформировавшейся в условиях моря с пониженной солёностью. Ранее в эту область входило и Аральское море, но в на-
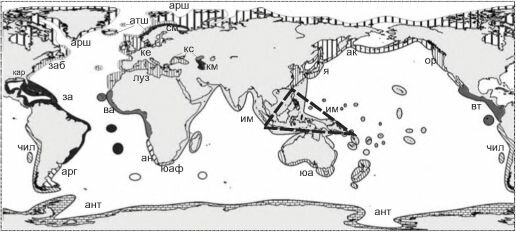
Рис. 6.11. Компилятивная схема биогеографического районирования шельфа. Аркто-бореальная область с провинциями и/или подобластями: арш — арктическая шельфовая, заб — западноатлантическая бореальная, кар — каролинская, атш — атлантическая шельфовая, см — скандинавская мелководная, ке — кельтская, луз — лузитанская, я — японская, ак — алеутско-камчатская, ор — орегонская. Тропическая атлантическая область: за — западно-атлантическая, ва — восточноатлантическая. Тропическая индо-тихоокеанская область: им — индо-малайская, вт — восточнотихоокеанская.
Нотальная (умеренная) область: чил — чилийская, арг — аргентинская, ан — анголо-намибийская, юаф — южноафриканская, юа — южноавстралийская.
Антарктическая область: ант — антарктическая.
Каспийская область: км — каспийская морская, кс — каспийская соло но вато водная.
стоящее время его аборигенная биота полностью вымерла и теперь его можно рассматривать скорее как часть средиземноморского региона.
Среди всех районов по таксономическому разнообразию резко выделяется Индо- Малайский регион. Так, моллюсков там обитает 6000 видов (из 27000, известных в Мировом океане), 800 видов иглокожих (из 6000), 4000 видов рыб (из 20000). Впрочем, это не удивительно. Мелководья этого района имеют площадь 6,5 млн. км2, тогда как всех остальных тропических регионов вместе взятых — только 2 млн. км2. К тому же, эти мелководья разбиты на большое количество более или менее изолированных участков, что приводит к частичной изоляции. Колебания уровня воды в кайнозое, несомненно, изменяли степень изоляции и связи разных его участков, как за счёт возникновения и устранения преград в виде суши, так и за счёт неизбежного изменения системы течений.
G.J. Vermeij (1978) приводит многочисленные примеры родов, обитавших ранее (судя по палеонтологическим данным) циркумтропически, а ныне являющихся палеоэндема- ми Индо-Малайского региона. Различия других тропических регионов также в значительной степени объясняются дифференцированным вымиранием. Имеется много причин, по которым таксоны, имевшие в прошлом пантропическое распространение, ныне отсутствуют в одном или нескольких регионах или, что наблюдается часто, ограничены в своём распространении Индо-Малайским регионом:
- Климатические изменения в разных районах тропиков были неодинаковыми, Индо-Малайский регион — единственный, в котором не было существенного изменения абиотических условий с момента его возникновения. Выше уже был приведён пример восточной тропической Пацифики, в котором в недавнее время вымерли коралловые рифы, головоногие моллюски внешней части шельфа, брахиоподы и исчезли луга морских трав. Аналогичные события были и в восточной тропической Атлантике, там также сейчас отсутствуют луга морских трав и брахиоподы. Существенное вымирание было и в Вест-Индии.
- Большая площадь Индо-Малайского региона определяет то, что в разных его районах условия весьма разнообразны, причём разные биотопы занимают в его пределах существенные площади. Это позволяет существовать разнообразным специфическим ассоциациям и ЦС.
Сам Индо-Малайский регион также неоднороден:
- Наибольшее число видов отмечено из его центра, показанного на карте треугольником, впервые выделенного J.C. Briggs. В пределах некоторых семейств и родов к этому треугольнику приурочены наиболее специализированные формы, тогда как генералисты обитают по его периферии.
- Число видов и родов мадрепоровых рифообразующих кораллов уменьшается по мере удаления от этого треугольника, в то время как средний возраст родов, судя по палеонтологическим данным, по мере удаления от этого треугольника растёт. Растёт и возраст Balanoida и стебельчатых морских лилий.
- Имеются ареалы с разрывом в треугольнике Индо-Малайского региона, где такие виды замещаются другими, вероятно, более молодыми.
J.C. Briggs (1995) полагает, что эти данные свидетельствуют о непрерывно идущем процессе видообразования в этом регионе. Много весьма интересных с общебиологической точки зрения вопросов может быть изучено здесь. К сожалению, он изучен крайне недостаточно. Например, число полихет, известных из Индо-Малайского архипелага, меньше, чем известных с шельфа Северного Ледовитого океана, хотя, судя по другим, лучше изученным группам бентоса, разница должна быть на порядок. Стремительное разрушение человеком коралловых рифов Индо-Малайского архипелага — проблема мирового масштаба. Эта проблема даже более значимая, чем уничтожение тропических лесов, поскольку рифы, в отличие от лесов, не только центр разнообразия, но и играют существенную роль в связывании С02 амосферы.
Имеющиеся данные показывают, что некоторые из мелководных таксонов, несомненно Индо-Малайского происхождения, оказались способными преодолеть биогеографические барьеры и заселили соседние регионы.
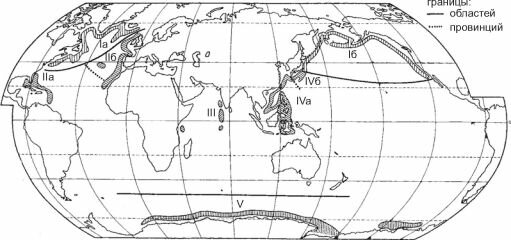
Рис. 6.12. Районирование батиали Мирового океана по брахиоподам О.Н. Зезиной (1973).
I — Бореальная область: 1а — Североатлантическая подобласть; 16 — Северотихоокеанская область; II — Амфи- атлантическая область: Па — Центральноатлантическая провинция; 116 — Лузитано-Мавританская провинция (переходного характера); III — Западно-индоокеанская область; IV — Западно-Тихоокеанская область: IVa — Малайская провинция; IV6 — Японская провинция (переходного характера); V — Антарктическая область.
Батиаль
Батиаль — наименее изученная часть Мирового океана. Это связано с высокой мозаичностью её сообществ, сложным рельефом, делающим недостаточно эффективными стандартные орудия лова (трал и дночерпатель) и существенным изменением абиотических условий в её пределах. Само существование единой вертикальной зоны «батиаль»
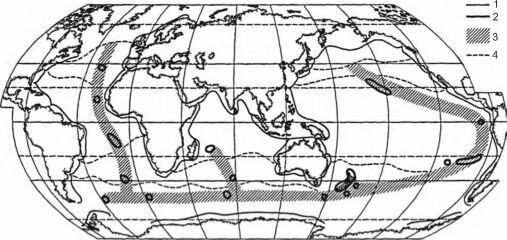
Рис. 6.13. Гипотетическая схема фаунистического районирования сублиторали и верхней батиали
Мирового океана А.Н. Миронова (1994).
- — фаунистические границы между приконтинентальными и океаническими регионами (по Миронову, 1989),
- — границы океанических экзотических регионов — регионов с изолированными островами и подводными горами, для которых свойственен повышенный эндемизм (островной эффект), 3 — зона расположения океанических экзотических регионов, которая одновременно служит пределом распространения для западных и восточных океанических таксонов, 4 — зимние (февраль в Северном полушарии и август в Южном) изотермы поверхностных вод 10 и 20° С (по Степанову, 1974).
следует считать пока ещё недостаточно обоснованным. Поэтому не удивительно, что было предложено всего несколько схем биогеографического районирования батиали Мирового океана, одна из них — О.Н. Зезиной (1973, рис. 6.12). К сожалению при построении схемы был использован индекс Престона, непригодный для целей районирования (подробнее — см. в предыдущей главе).
А.Н. Миронов (1989, 1994) предложил единую схему районирования сублиторали и батиали, включая талассобатиаль (рис. 6.13). Согласно ей сублиторальные и батиальные районы имеют совпадающие широтные фаунистические границы, положение которых, как он предположил, приурочено к изотермам.
Также он показал, что для районов талассобатиали, особенно изолированных океанических островов (эти районы он назвал «экзотические») характерно наличие большого числа эндемов, как и следовало ожидать из их островного положения (см. предыдущие главы). Дальнейшие исследования должны также выявить и остальные черты островных биот (уменьшенное общее число видов, букетное видообразование и т. д.). Такие районы приурочены преимущественно к срединно-океаническим хребтам, а также к цепочкам островов и подводных вулканов, образованных в результате действия горячих точек. Некоторые эндемы широко распространены вдоль этих цепочек островов и подводных гор. Одновременно А.Н. Миронов показал, что эти же районы являются границами для распространения батиальных видов с широкими ареалами, приуроченными к восточным или западным частям океанов.
Еще по теме Шельф:
- Фронт края шельфа.
- Биогеографическое районирование
- Прибрежный фронт.
- 2. Функционирование биоценозов
- Пограничный эффект
- Гетеротрофные сообщества
- Экономическое значение рыб
- Метод значимости биогеографических границ
- Аркто-бореальное
- Взаимодействие морских бореальных биот Атлантики и Пацифики
- Вертикальная зональность