ПРОДВИЖЕНИЕ ЯЙЦЕВОЙ КЛЕТКИ
Эффективность осеменения зависит не только от спермия, но и от судьбы яйцеклетки после ее выделения из фолликула. В момент овуляции яйцевая клетка, окруженная лучистой короной (клетки фолликулярного эпителия), вместе с фолликулярной жидкостью выделяется в брюшную полость (рис.
39).
Вследствие тесной связи яйцепровода с яичником яйцевая клетка обычно попадает на ответвления бахромки. Этому же способствуют помимо анатомических условий динамические свойства бахромки и яйцепровода. Так, у обезьян к моменту овуляции сокращаются мышцы яйцепровода и маточной брыжейки.
гут частично выделяться и через ее половую систему. Дача внутрь свинца, никотина, фосфора сильно сокращает время переживаемости спермиев и в 50 % случаев вызывает аборт. Мышьяк, ртуть, коллоидальное серебро и железо несколько укорачивают срок жизни спермиев в матке.
Само по себе поступление спермы в половые пути самки влечет ответную реакцию организма, выражающуюся в гиперемии половых органов, образовании спермиолизинов, спермиотокси- нов, спермиоагглютининов.
Спермии самца одного вида, введенные в матку самки другого вида, погибают быстрее по сравнению со спермиями, введенными в матку самки от самца того же вида.
Результаты экспериментальных исследований и большое количество практических наблюдений за эффективностью осеменения в зависимости от интервала между осеменением и овуляцией позволяют считать, что оплодотворяющая способность спермиев в половом аппарате самок млекопитающих сохраняется максимум в течение 36— 48 ч. Из учета этих сроков и следует исходить при выборе времени для осеменения животных.
Бахромка вследствие наполнения ее сосудов кровью становится упругой и охватывает яичник, как бы подготавливая ложе для выделяющегося яйца.
У кобылы, свиньи, собаки, самки медведя, хорька и у некоторых других животных яичник заключен в бахромку, или складку, яйцепровода; таким образом создаются такие взаимоотношения, при которых выделившееся яйцо, как правило, должно попасть непосредственно на бахромку.
У других животных (корова, овца, коза) бахромка развита сравнительно слабо и не может охватить овуляционный участок яичника, поэтому нельзя объяснить проникнове-
ние яйца в яицепровод исключительно эректильным состоянием бахромки. Процесс этот некоторые авторы объясняют токами жидкости из брюшной полости в матку. Возбудителем этих токов В. С. Груздев считает колебания ресничек эпителия бахромки и яйцепрово- да. Мерцательные движения их, направленные в полость матки, перегоняют жидкость из брюшной полости в матку, а образовавшиеся токи увлекают выпавшую в брюшную полость яйцевую клетку. Такого же мнения придерживается К. К. Скробанский, который объясняет возникновение токов жидкости еще и сокращением брюшного пресса, повышающим внутрибрюшное давление в момент овуляции.
По наблюдениям медиков, в большинстве случаев трубной беременности яйцевая клетка выделяется правым яичником , а плод развивается в левой трубе. Это обстоятельство позволило предположить, что после овуляции яйцевая клетка может «странствовать» по брюшной полости, а затем попадать в тот или иной яйцепровод. Специальные опыты послужили основанием к введению понятий о наружном и внутреннем «странствовании» яйца или зиготы (migratio externa et migratio interna). Первое заключается в том, что яйцевая клетка по выделении ее из яичника некоторое время плавает в капиллярных щелях между петлями кишечника и лишь позднее проникает в полость матки через просвет одной из труб. Внутреннее «странствование» выражается в перемещении зиготы (возможно, и зародыша) из одного рога в другой через тело матки и может быть объяснено сокращением мышцы матки.
Сроки продвижения яйцевых клеток по яйцепроводам, по-видимому, могут сильно варьировать. Специальные опыты и клинические наблюдения позволяют допустить, что прохождение яиц по яйцепроводам у всех животных продолжается 1—3 сут. У коров яйцо в течение
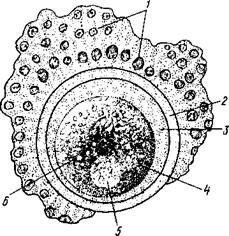
![Рис.<div class=]()
39. Схема строения яйца:" />
Рис. 39. Схема строения яйца:
1 — клетки лучистого венца; 2— прозрачная оболочка; 3 — перивителлиновое пространство; 4 — цитоплазма; 5 — ядро; 6— желточные зерна
первых 6—12 ч после овуляции проходит треть яйцепровода. Дальнейшее продвижение яйцеклетки происходит медленно — примерно около 0,1см в 1
ч. После овуляции яйцеклетка способна к оплодотворению в течение 4—6 ч.
У птиц этот срок составляет всего только 20—25 мин (А. Д. Курбатов и др.). Если оплодотворения не произошло, яйцо погибает и рассасывается. Время жизни яйцеклеток у некоторых животных и поступление зигот в матку приведены в таблице 12. 12.
Сроки жизни яйцевых клеток, продвижения зигот и имплантации зародышей (по Р. X. Хантеру) Корова 10-12 72-84 9-11 22 Овца 10-15 66-72 7-8 15 Свинья 8-12 46-48 6 18 Лошадь 8-10 140-144 8 37 Вид
живот
ного
Максимальное время жизни яйца после овуляции, ч
Оплодотворение — физиологический процесс, заключающийся в слиянии яйца и спермиев с последующей их ассимиляцией и диссимиляцией, в результате чего образуется новая клетка (зигота), обладающая двойной наследственностью.
В 1651 г. У. Гарвей высказал свое историческое изречение: «Все новое — из яйца», породившее направление овистов (ovo — яйцо).
После открытия спермиев студентом Гаммом и его учителем А. Г. Левенгуком (1677 г.) в биологии стала господствовать теория преформизма, утверждавшая, что спермий, являясь микроскопически малым, но вполне сформированным организмом (animalculum — зверек, homun- culum — человек), попадает во время полового акта в полость матки и использует материнский организм как почву для своего роста (не развития!). Поэтому в русской литературе еще встречаются два порочных термина для обозначения спермиев: сперматозоид (семя, подобное животному) и живчик (живой человечек).
В 1759 г. русский академик К. Ф. Вольф опубликовал свой труд «Теория зарождения», которым положил начало теории эпигенеза, т.
е. теории развития. С этого времени началась двухвековая борьба сторонников двух течений: эпигенеза и преформизма.
В 1827 г. русский ученый К. Бэр открыл яйцевую клетку млекопитающих и заложил основы современной эмбриологии. После открытия яйцеклетки одни преформисты усматривали готовый микроорганизм в спермии, другие — в яйце.
В 1876 г. Р. Гертвиг и Фоль впервые наблюдали проникновение спермия в яйцевую клетку морского ежа. Наука стала приближаться к пониманию сущности оплодотворения.
Во время оплодотворения происходят взаимная ассимиляция (assimilatio — взаимопоглощение) и диссимиляция (dissimilatio — расподобление) яйца и спермия, в результате которых уже не существует ни яйца, ни спермия, а образуется новая, третья клетка — зигота (от греч. zygotos — соединенный вместе), не тождественная ее двум первоисточникам. Поэтому широко применяемое в биологии называние зиготы оплодотворенным яйцом не соответствует действительности.
Сложный процесс оплодотворения происходит в верхней трети яйцепрово- да. Спермии быка непосредственно перед оплодотворением должны не менее 6
ч находиться в абдоминальной части яйцепровода. За это время они «созревают» и становятся способными оплодотворять. Такое явление называется капа- цитацией (инкубацией) спермы. У спермиев барана капацитация длится 1— 1,5
ч, а у хряка —2—3 ч. Оплодотворение может произойти только при достаточном скоплении спермиев. Количество их значительно колеблется даже у одного и того же вида животных. Спермии ожидают яйцеклетку. Последняя, как и спермии, должна «дозреть». Этот процесс также происходит в яйцепрово- де, куда яйцеклетка попадает в стадии овоцита. Там она при наличии активных спермиев выделяет направительные тельца и созревает. Сколько времени требуется для «дозревания» яйцеклетки, пока неясно. У овец, видимо, не более 2 ч.
Установлено, что для нормального процесса оплодотворения, а стало быть, получения жизнеспособного плода необходимо соединение только полноценных половых клеток.
Неполноценные гаметы не вступают в процесс оплодотворения или вступают, но зародыши гибнут на ранней стадии. В связи с этим важное значение имеют время осеменения, возраст половых клеток и активность спермиев.
Оплодотворение у животных состоит из нескольких стадий.
Первая стадия — денудация — характеризуется тем, что яйцеклетка, проходя по яйцепроводу, освобождается от окружающих ее фолликулярных клеток (лучистого венца). Основная роль в денудации принадлежит спермиям, которые, внедряясь между клетками фолликулярного эпителия, выделяют ферменты, главным образом гиалуронидазу. За счет этих ферментов и неизвестных еще факторов разжижается гиалуроновая кислота, входящая в состав студенистого вещества, связывающего клетки лучистого венца. Рассеивание клеток лучистого венца — это не видовая особенность, и оно может происходить под влиянием спермиев животных другого вида. Так, при осеменении свиней смесью спермы быка и хряка спермии быка освобождают яйцо от фолликулярных клеток, а проникают в него только спермии хряка. Процессу денудации способствуют также механические препятствия, обусловленные ворсинками слизистой оболочки яйцепровода.
Для оплодотворения не обязательно полное освобождение яйцеклетки от клеток лучистого венца. Достаточно лишь места, чтобы спермии проникли через прозрачную оболочку яйцеклетки в околожелточное пространство. Этот процесс, составляющий вторую стадию оплодотворения, в видовом отношении более специфичен. В это время заканчивается созревание яйцеклетки (выделение второго направительного тельца). Ядро, содержащее гаплоидное число хромосом, превращается в женский пронуклеус.
На третьей стадии оплодотворения один, реже несколько спермиев проникают через желточную оболочку яйцеклетки в ее цитоплазму. Это уже строго специфический процесс, поскольку в яйцеклетку могут проникать только спермии своего вида. При этом проникает не весь спермий, а лишь его головка и шейка. Внедрившись в цитоплазму яйцеклетки, спермий претерпевает большие изменения.
Головка спермия быстро увеличивается в размере в десятки раз; вследствие ассимиляции цитоплазмы яйца она достигает величины ядра яйцеклетки и превращается в мужской пронуклеус, имеющий, как и женский пронуклеус, половинный набор хромосом.
На четвертой стадии пронуклеусы (ядра яйцеклетки и спермия) постепенно сближаются, вступают в тесный контакт, быстро уменьшаются в объеме и полностью сливаются. Этот процесс, основной в оплодотворении, является еще недостаточно изученным. Образуется качественно новая клетка (зигота), ядро которой содержит диплоидное число хромосом. Образовавшаяся зигота, таким образом, имеет двойную наследственность. Она получает могучий стимул для дальнейшего развития и начинает быстро дробиться (рис. 40). После первого деления зиготы образуются два бластомера (дочерние клетки); оба бластомера дают четыре внучатых, а четыре внучатых — восемь правнучатых бластомеров и т. д. У млекопитающих такой закономерности нет: количество бластомеров у них даже в начальных стадиях не всегда бывает четным. В отличие от деления одноклеточных организмов клетки, образующиеся в результате деления зиготы, не расходятся, а остаются соединенными; в первый период общая величина зародыша не изменяется, поэтому деление зиготы называется дроблением. Дробление зиготы коровы длится около 8 сут. В течение первых 4 сут дробление происходит в яйцепро- воде, а затем в одном из рогов матки. Уже через 48 ч после овуляции зигота состоит из двух бластомеров, через 72 ч — из трех, через 84 ч — из шести и через 96 ч она поступает в стадии мору- лы в рог матки. Зигота по размеру равна яйцевой клетке или незначительно превышает ее, так как с каждым дроблением при увеличении количества бластомеров величина каждого из них соответственно уменьшается.
Наружный слой бластомеров, прилегающий к прозрачной оболочке, называется трофобластом (питающим листком); бластомеры, заключенные в трофобласт, образуют э м б - риобласт (зародышевыйлисток).
Полиспермия (переоплодотворение). Проникновение в цитоплазму яйцевой клетки двух или нескольких спермиев называется полиспермией. Из сотен миллионов спермиев, попадающих в половые пути самки при коитусе, лишь тысячи достигают бахромки.
В норме после проникновения в яйцо одного спермия другие не пропускаются в силу так называемой зональной реакции. У стареющих яиц зональная реакция нарушается, что обусловливает невозможность проникновения спермия в яйцо, а иногда под влиянием каких-то условий происходит слияние ядра яйца с несколькими спермиями. Это происходит, вероятно, тогда, когда приблизившиеся к яйцу спермии одновременно проникают внутрь его.
Полиспермия нарушает течение эмбриогене
за, приводит зиготу к гибели или, по мнению некоторых авторов, обусловливает развитие уродливых плодов (паразитизм, дицефализм и др.).
При физиологической полиспермии у млекопитающих спермии, не сливающиеся с ядром, по- видимому, только активизируют зиготу к дроблению аналогично реакции организма на введение инородного белка.
Множественное оплодотворение, или суперфекундация (вирегГесипскПо). Это оплодотворение нескольких яйцевых клеток в период одного полового цикла, но спермой разных производителей. Оно часто наблюдается у многоплодных животных (собаки, свиньи, кошки), реже — у кобыл и коров. Суперфекундация, развившаяся в результате осеменения производителями, принадлежащими к одной породе и одинаковой с самкой масти, часто остается незамеченной; она выявляется, когда несколько производителей, принимавших участие в осеменении самки, принадлежит к разным породам и даже видам. Так, из- за недосмотра в стадию возбуждения у породистых собак наряду с чистопородными могут родиться щенки с признаками двух или трех пород, нескольких мастей и т. д. Отмечены случаи суперфе- кундации у свиней, когда рождаются поросята различных мастей; у кобыл, когда одновременно рождаются нормально развитые жеребенок и муленок.
В настоящее время в животноводческой практике суперфекундацию применяют для повышения оплодотворяемое- ти и жизненности приплода. С этой целью при естественном осеменении практикуется коитус с двумя самцами с интервалом 10—15 мин.