Каспийское море
Каспийское море — самый маленький регион ранга области в предлагаемой биогео- графической схеме. Выше уже говорилось о причинах формирования здесь высокоэндемичной биоты, поэтому остановлюсь только на биогеографической структуре региона.
Само Каспийское море занято двумя ЦС: одна система занимает Северный Каспий, другая — Средний и Южный (см. главу 1). Своеобразна вертикальная структура Среднего и Южного Каспия. В отличие от всех других областей Мирового океана и даже многих крупных пресноводных водоёмов, здесь нет вертикальных зон. Бентос Каспия обитает преимущественно на глубинах менее 50-100 м и в основной своей массе не спускается глубже, т. е. это — типичный верхнесублиторальный бентос, что довольно обычно, по крайне мере в бореальных водах Атлантики и Дальнего Востока, где на этих глубинах также проходят границы между верхнє- и нижнесублиторальными биогеографическими регионами, на которых кардинально меняется видовой состав. Необычность Каспия состоит в том, что в нём этой смены не происходит: нижнесублиторальной фауны нет, нет и соответствующей ЦС, из всех известных видов лишь один, Didacna profundicola, обитающий на глубинах 75-409 м (Атлас ..., 1968; Карпинский, в печати) характерен именно для элиторали. Эту давно известную особенность фауны наиболее глубоководных районов Каспия объясняют плохим кислородным режимом (Зенкевич, 1963) или недостатком пищи (Карпинский, 2002). Но эти причины могут объяснять только вертикальное распределение видов, а не полное отсутствие специфической нижнесублиторальной фауны. Отсутствие такой фауны можно понять при интерпретации истории биоты Каспия с позиций парадигмы структурализма.Как было показано в предыдущих главах, новая ЦС возникает всякий раз, когда появляется достаточно обширный длительно существующий биотоп, который не может быть заселён существующими ЦС. Поскольку глубины Каспия не населены существующими ЦС, а своей ЦС там не возникло, то или биотоп недостаточно большой, или время его существования невелико.
Первая причина кажется маловероятной, поскольку свои ЦС (или, что то же самое, — биогеографические регионы) с высокоэндемичной фауной возникают в пресноводных озерах гораздо меньшего размера. По-видимому, в Каспии глубоководная ЦС не возникла по второй причине — из-за кратковременности существования данного биотопа. Это заставляет предположить, что в Каспии в норме часто возникали и возникают заморы на глубинах, что препятствует образованию нижнесублиторальной ЦС. Посмотрим как это предположение соотносится с фактами.Аральское, Каспийское, Азовское и Чёрное моря, а также озеро Бива на Балканах являются остатками Паратетиса — внутриконтинентального моря, в периоды максимального развития простиравшегося между 40° и 50° с. ш. от Альп до Тянь-Шаня и превышавшего по площади современное Средиземное море не менее, чем в два раза. Паратетис обособился от Тетиса в олигоцене в результате возникновения гор Альпийского пояса (Альпы, Балканы, Крымские и Кавказские горы) (рис. 1.30). Изменения состава биоты Паратетиса в первую очередь определяли изменения солёности, зависящие от степени его связи с Тетисом, а после исчезновения Тетиса — со Средиземным морем. Лишь в самый последний период устанавливалась временная связь с распреснёнными районами Северного Ледовитого океана.
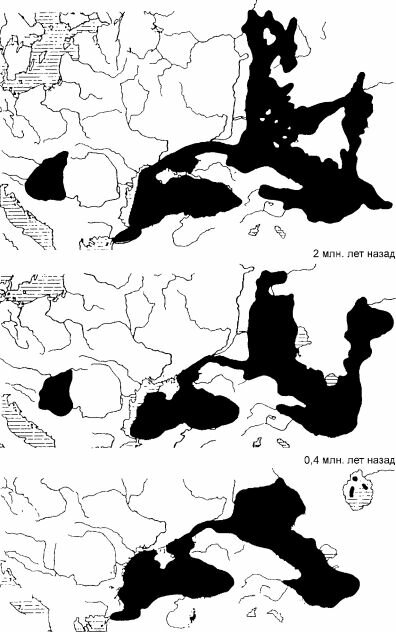
Поскольку восточная часть Паратетиса — один из основных нефтегазоносных районов СССР, его истории посвящено огромное число работ, и последовательность событий известна в деталях. В целом эта история — череда распадов востока Паратетиса на водоёмы в разной степени изолированные (рис. 6.18) и их объединения. В общем изоляция приводила к снижению солёности, порой существенному, тогда как установление связи с Тетисом или Средиземным морем — к увеличению солёности до близкой к нормальной океанической. Биота современного Чёрного моря, за исключением эстуарных районов, яв-
Рис. 6.18. Паратетис.
По И.В. Аладину и др
(2004) ч 'Л.
3 млн. лет назад
ляется недавней (возраст примерно 7150 лет) производной биоты Средиземного моря, из состава которой полностью или почти полностью выпали стенога- линные таксоны (иглокожие, мшанки, брахиоподы, оболочники и др.), а число видов остальных таксонов гораздо меньше того, которое можно было бы ожидать, имей Чёрное море нормальную солёность.
Казалось бы, и остальные водоёмы, относящиеся к Каспийской области, для которых характерна пониженная солёность, должна населять биота сходного состава, с более или менее сильным эндемизмом, причём эндемы должны происходить от немногих пред- ковых форм. Действительно, в ней отсутствуют стеногалинные группы, а биота происходит от немногих предковых форм — ситуация довольно банальная, причины которой подробно рассмотрены в главе 4. Но эти факты мало что дают для проверки нашей гипотезы. Гораздо интереснее другое: кроме стеногалинных таксонов в составе биоты Каспия отсутствуют и таксоны, которые по современным абиотическим условиям, казалось бы, могли вполне входить в её состав и, более того, — успешно существуют в настоящее время, после того, как они были перенесены в Каспий человеком. Эти особенности состава Каспийской области невозможно объяснить только пониженной солёностью или несколько иным солевым составом.
Неогеновые бассейны Паратетиса с нормальной солёностью были населены сообществами бентоса, в составе которых основную роль играли моллюски — двустворчатые и гастроподы, роль полихет из-за плохой сохранности в ископаемом виде, неясна, возможно, что она также была существенна. Трофическая зональность была характерной для современных морей: от берега к открытому морю располагались зоны фильтраторов эпифауны твёрдых грунтов, фильтраторов инфауны и собирающих детритофагов, среди которых были и моллюски рода АЪга. К последним в качестве сопровождающих форм, присоединялись способные плавать фильтраторы эпифауны (двустворчатые моллюски-гребешки) и в небольшом количестве фильтраторы инфауны (Невесская, 1999).
В водоёмах с солёностью существенно ниже океанической сообщества моллюсков сотояли только из фильтраторов (инфауны и эпифауны твёрдых грунтов). Именно этот состав и характерен для автохтонов Каспийской области: двустворчатые моллюски представлены только фильтраторами инфауны (Cardidae) и эпифауны твёрдых грунтов (Dreis- senidae). Т. е.
с понижением солёности из состава биоты выпадали моллюски-детритофа- ги. Успешная акклиматизация АЪга, виды которой имелись в исходной биоте, но отсутствуют в современной каспийской, показывает, что выпадение моллюсков-детритофагов необъяснимо лишь изменением солёности. По-видимому, существовали какие-то причины, мешавшие развитию видов моллюсков-детритофагов и приводившие к вымиранию уже возникших.Биотоп детритофагов — это мягкие грунты с большим содержанием органики и пониженной гидродинамикой. И то, и другое ведёт к снижению концентрации кислорода в придонной воде. Солёность водоёмов Паратетиса часто (как и сейчас) снижалась ниже 24,695%о — солёности, ниже которой вертикальная циркуляция начинает идти по типу пресных водоёмов (см. главу 1). Дополнительно вертикальную циркуляцию снижал боль
шой пресноводный сток, создававший рас- преснённую линзу на поверхности. Всё это приводило к ухудшению аэрации придонных слоёв, и, по-видимому, возникновению частных заморов. Экологическая ниша моллюсков-детритофагов при появлении подходящих условий (улучшении кислородного режима) заполнялась автохтонными дериватами, например двустворчатыми моллюсками Limnocardiinae и Dreissenidae (рис. 6.19), но через некоторое время они вымирали. Поэтому биотопы, занятые в водоёмах с океанической солёностью сообществами моллюсков-детритофагов, в Каспийской области заняты сообществами, в которые входят мол- люски-фильтраторы (эпифауны твёрдых грунтов и инфауны), а в автохтонной биоте Каспия детритофаги представлены ракообразными и полихетами Ampharetidae. Двустворчатые моллюски же сравнительно мало специализированы, т. е. обладают широким спектром питания: от детритофагии до фильтрации. Аналогичная ситуация существовала и в водоёмах предшествовавших Каспию, формировавшихся на территории Паратетиса, например, в позднесарматском и раннеакчагыльском водоёмах. Сходным образом из состава бентоса выпали бентосные хищники (гастроподы, декаподы) (Невесская, 1999).
В автохтонной биоте Каспия хищные животные (включая бентофагов) принадлежат почти исключительно к рыбам (осетровые, бычки), кроме них в Каспии широко распространён пресноводный по происхождению рак Astacus pachypus. Недавно вселившийся краб Rhithropanopeus har- risii широко распространился по Каспию, что также свидетельствует о недоиспользован- ности ресурсов хищниками.Итак, история Каспийского моря и Паратетиса в целом однозначно свидетельствует о том, что, кроме колебаний солёности, существенным фактором, влиявшим на формирование биоты, было регулярное возникновение заморов. И действительно, в котловинах Среднего и Южного Каспия в XX в. регистрировали наличие сероводорода. Таким образом, часто возникавшие заморы оказали существенное влияние даже на состав биоты верхней сублиторали. Логично допустить, что глубже верхней сублиторали — в биотопе нижнесублиторальной биоты — концентрация кислорода была ещё ниже, что не позволило сформироваться нижнесублиторальной биоте. Трофический фактор и биотические взаимоотношения, которые М.Г. Карпинский (в печати) считает также существенными, на этом фоне, вряд ли сыграли сколь-нибудь значимую роль.
Еще по теме Каспийское море:
- Каспийско-черноморские сельди
- УТОНУВШИЕ В МОРЕ ЛИСТВЫ
- ГЛАВА ТРЕТЬЯ НА ЗЕМЛЕ, В НЕБЕСАХ И НА МОРЕ
- Формы акклиматизации.
- Черепахи
- Гренландский кит Balaena mysticetus Linnaeus, 1758 (I, II, VI, 268)
- ДЕЛЬфИН-БЕЛОБОЧКА И АфАЛИНАDelphinus delphis L., Tursiops truncatus Montagu
- Черноморская афалина Tursiops truncatus ponticus Barabasch, 1940 (II, 251)
- 18.1. БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ И СОСТАВ ПЧЕЛИНОЙ СЕМЬИ
- КЛАСС КРУГЛОРОТЫЕ — CYCLOSTOMATA
- Влияние морских течений на распределение температу р в водных массах.
- Проверка двух теорий эволюции
- Образование льда в морской воде
- Атерйновые (Atherinidae)