Глава 7. ЭТОЛОГИЧЕСКАЯ СТРУКТУРА И СОЦИАЛЬНАЯ ОРГАНИЗАЦИЯ
Обитание сайгаков в открытых ландшафтах аридных зон определяет наличие у этих копытных специфичной отологической структуры, при которой формирование определенных группировок особей, различных по величине и t составу, является важнейшей адаптационной особенностью популяций.
Стадный образ жизни копытных формируется на основе сложно-рефлекторных отношений между особями данного вида. В основе социальной организации населения животных лежит взаимное тяготение друг к другу особей одного вида, которое присуще многим копытным открытых пространств (Насимович, 1955).Под стадом у копытных обычно понимается определенная группа животных, размер которой колеблется от нескольких особей до тысячных табунов, ведущих совместную и взаимосвязанную жизнь в какой-то определенный период времени. Подобные группы копытных обычно действуют как единое целое, т.е. все особи этой группы совместно делают переходы, пасутся, отдыхают, а также все вместе защищаются от врагов. Таким образом, основным показателем стада копытных следует считать общность форм поведения особей.
Стадо - определенную группировку особей - почти всегда можно достаточно четко дифференцировать, поскольку даже широко рассредоточенные пасущиеся сайгаки все-таки держатся определенной группой, где отдельные особи не уходят далеко друг от друга, но и соблюдают индивидуальную дистанцию. При приближении опасности все особи такой группы сначала собираются в четко обособленное стадо, которое и уходит от преследователя в одном направлении.
Взаимное тяготение отдельных особей сайгаков друг к другу четко прослеживается на размерах индивидуальной дистанции, которая для этого вида в летний период составляет: на отдыхе - 2,7 + 0,5 м; при бегстве - 0,3 + 0,1 м; при пастьбе - 5,0 + 1,4 м (Баскин, 1976). Однако стадо у многих копытных не является какой-то неизменной и постоянной группой одних и тех же особей.
Очень часто можно наблюдать, как одно убегающее стадо соединяется с другим или наоборот, большое стадо может разделиться на отдельные, более малочисленные группы особей. *Такая неустойчивость по составу - характерная особенность стадных группировок сайгаков (Жирнов, 1961; Баскин, 1976). Тем не менее, несмотря на непрочность связей между конкретными особями, сайгаки как вид могут нормально существовать только в определенных агрегациях - стадах. Более того, в определенные периоды года сайгаки существуют и в более крупных группировках - в скоплениях, которые являются результатом концентрации отдельных стад на определенных участках территории поблизости одно от другого. В других скоплениях, например, во время миграций, а также на местах рождения молодняка - в "родильных домах" - трудно выделить отдельные стада, и такие агрегации формируются под воздействием одного доминирующего фактора - в результате локализации на удобных для родов участках местности или на основе сходства активности во время сезонных миграций или других массовых перемещений сайгаков. Мы не согласны с Л.Баскиным (1976) в том, что в скопле-
ниях сайгаков "нет и сходства в активности, ни единого направления движения". В скоплениях мигрирующих животных, а также в "родильных домах" четко выделяются признаки взаимосвязанности многих особейнаправление перемещений и периодичность активности (пастьба, кормление молодняка и т.д.). Следует отметить, что вопросы взаимоотношения между особями у высокостадных копытных, в частности, у сайгаков еще изучены недостаточно полно. В частности, не использовались методы мечения животных при изучении поведения этих ЖИВОТНЫХ. \
Непрочность связей между отдельными особями внутристадных группировок, по нашему мнению, является важнейшей экологической особенностью таких высокоподрижных животных как сайгаки, поскольку номадизм исключает постоянство состава внутрипопуляционных группировок. Действительно, наиболее постоянные и стойкие группы, состоящие из самки с 1-2 молодыми.
Такие группы сохраняются на период лактации, т.е. с мая по август -сентябрь. Устойчивость самок и молодых определяется необходимостью выкармливания молодняка молоком. В литературе имеются указания, что такие семьи сохраняются до гона (Васенко, 1950 и др.), но прямые, строго документированные наблюдения, например с применением мечения, отсутствуют. Другой пример прочных связей внутри группировок сайгаков относится к периоду гона, когда по крайней мере на несколько дней взрослый самец - владелец гарема, выполняющий роль лидера - вожака, сохраняет определенную группу самок на своем "гаремном" участке. Постоянство таких групп активно поддерживается доминирующим самцом путем подавления подвижности гаремных самок, которые даже и в этих условиях стремятся покинуть группу (Фандеев, 1961 и др.). В этом случае постоянство групп определенным образом связано с локализацией гаремных стад на определенной территории, т.е. с ограничением подвижности особей. *'• ' А‘Многолетними исследованиями в Северо-Западном Прикаспии и в Казахстане было достатолчно четко показано изменение в распределении встреч сгад резкой величины и состава по сезонам года, а также в зависимости от погодно-климатических и кормовых условий по годам (Жирнов, 1961; Фадеев, Слудский, 1982).
Изменение величины стада обычно принято выражать "показателем стадности" (средняя арифметическая величина, получаемая от общего числа особей во всех встреченных стадах за тот или иной период). При рассмотрении сезонных изменений групповой организации сайгаков подобный показатель не будет отражать истинного положения, поскольку размеры стад сайгаков в любой сезон года колеблются от маленьких групп до сотенных и тысячных табунов. В связи с этим при характеристике стадности нами выделялись следующие категории, характеризующие величину стада: 1) от 1 до 5 особей; 2) 6 - 10; 3) 11 • 19; 4) 20 - 49; 5) 50 - 100; 6) 101 - 200; 7) 200 - 500; 8) 501 - 1000; 9) свыше 1000 особей.
Как было установлено для многих видов копытных животных (Насимович, 1955 и др.), величина и состав стад существенно меняются в разные сезоны года, что определяется поведением животных и состоянием их среды обитания. Все имеющиеся материалы рассмотрим по сезонам года.
Р и с. 44. Встречаемость стад различных категорий с 1 по 10 (1), с 10 по 25 (2) и в конце
декабря (3)
Зима. Частота встреч разных разных стад сайгаков существенно меняется от декабря к январю. Если в декабре среди встреченных стад преобладают (61%) мелкие группы, состоящие из 1-20 особей, то в январе сайгаки держались более крупными стадами. В январе мелкие стада составляли всего 19% от всех встреч, и наиболее часто встречались средние по величине стада (56%); крупные и очень крупные отмечались также довольно часто (25%). В декабре встречи крупных стад были редки (5,5%), а средние составляли 34%.
Эти отличия характера стадности в декабре и январе-феврале обусловлены тем, что в декабре проходит гон сайгаков. С началом гона крупные стада разбиваются на мелкие группы. Сам процесс формирования гаремов, состоящих из самок, охраняемых взрослым самцом, происходит постепенно. Еще в первой декаде декабря нередко можно встретить крупные стада, состоящие из самок и самцов, численностью до 600 особей. В этот период также встречаются стада, состоящие из 50-100 голов (22,7%). В разгар гона, с 10 по 25 декабря, стада сайгаков представлены: 1) одиночками и мелкими группами, состоящими из взрослых самцов-одиночек и самцов-сеголеток (44,5%), гаремами из самок, охраняемых одним взрослым самцом (25%). Стада, численностью более 50 голов, отмеченные в конце декабря, представляли собой уже объединенные гаремные группы, у которых гон закончился. Неполовозрелые самцы в период гона формируют отдельные стада.
'/о
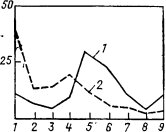
Рис. 45. Встречаемость стад различных категорий: 1 - в декабре; 2 - в январе
По наблюдениям в Казахстане социальная организация в зимний период была представлена следующими стадами. Группы численностью до 50 особей составляли в декабре 80%, от 51 до 100 - 14,9%, более крупные (в основном до начала гона) - 4,5%. После окончания гона гаремы объединяются, и в январе-феврале стада до 50 особей составляют 28,7-33,1%, тогда как в 51-500 особей •уже 56,5-59,3%, а свыше 500 - 10,4-12,0% (Фадеев, Слудский, 1982). Состав стад чаще смешанный, хотя встречаются отдельные небольшие группы и одиночки из числа ослабевших самцов, участвующих в гоне. Укрупнение стад происходит под влиянием погодных условий - сильного ветра, снегопадов, когда сайгаки быстро группируются и оставляют этот район. Видамо, это явление можно рассматривать как адаптацию, направленную на облегчение передвижения и добычи корма в условиях многоснежья.
Формирование чисто самцовых стад, отмеченное для Казахстана (Соколов, 1951), видимо, происходило при плохом урожае трав на зимних пасг-

Рис. 46. Отдельное стадо, состоящее из самцов. Калмыкия, май 1970 г.
Фото Л. В. Жирнова
Относительно состава стад в зимний период после гона мнения в литературе противоречивы. Одни авторы (Васенко, 1950) считают, что состав стад остается смешанным, тогда как другие (Соколов, 1951) указывают на полное обособление самцов, сильно истощенных гоном. Нам приходилось встречать смешанные стада как в конце декабря, так и в январе. Одновременно встречались мелкие группы только самцов и самцы-одиночки. Встреча таких самцовых групп указывает на тенденцию к обособлению, по-видимому, наиболее ослабевших самцов, участвовавших в гоне.
бюцах в связи с засухой. Для Прикаспия также известны случаи, когда зимой отмечались большие самцовые стада. Последнее имело место, например, в январе-феврале 1954 года в результате выпадения глубокого снега. За р. Кумой н в Ногайских степях в это время наблюдались стада, состоящие исключительно из взрослых самцов (Рак, 1956).
Формирование дифференцированных по полу стад определяется разной подвижностью самок и самцов. По этому признаку и происходит объединение особей в стада. Так, многоснежье сильно затрудняет передвижение животных, в результате и возникают отдельные стада крупных взрослых самцов.
Еще по теме Глава 7. ЭТОЛОГИЧЕСКАЯ СТРУКТУРА И СОЦИАЛЬНАЯ ОРГАНИЗАЦИЯ:
- Глава 4. Социальное поведение
- 2.11.1. Основные направления этологических исследований
- §1. Понятие о социальном поведении животных
- Глава 5 Пространственная организация популяций
- 7.5. «Социальные знания» и жизнь в сообществе
- Глава 14. СТРУКТУРА ПОПУЛЯЦИИ
- Глава 2 ОРГАНИЗАЦИЯ И ТЕХНОЛОГИЯ ОСЕМЕНЕНИЯ ЖИВОТНЫХ
- Глава 2 ОРГАНИЗАЦИЯ И ТЕХНОЛОГИЯ ОСЕМЕНЕНИЯ ЖИВОТНЫХ
- ГЛАВА 4 Организация жизни и ее основные характеристик
- Глава 1 Популяционный уровень организации живой материи
- ГЛАВА 3 СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА
- Глава 6 Структура популяций
- Глава У. СТРУКТУРА И СОСТОЯНИЕ ПОПУЛЯЦИЙ ЛОСЯ
- ГЛАВА 3 ЕДИНСТВО МАТЕРИАЛЬНОЙ СТРУКТУРЫ МИРА
- Глава 9. СТРУКТУР А ПОПУЛЯЦИЙ (ПОЛОВОЙ И ВОЗРАСТНОЙ СОСТАВ)
- ГЛАВА 10 БИОЛОГИЧЕСКИЙ ВИД. ПОПУЛЯЦИОННАЯ СТРУКТУРА ВИДА
- Глава V О ВЛИЯНИИ ПАРАЗИТНОГО ОБРАЗА ЖИЗНИ НА ЭВОЛЮЦИЮ ПОЛОВЫХ СТРУКТУР
- 8.8.2. Вклад социальной и биологической компонент в общую смертность в историческом времени и в разных популяциях
- Организация биосферы
- Международные фе-линологические организации