ХИМИЧЕСКИЙ АСПЕКТ ИЗУЧЕНИЯ ДЕТЕРМИНАЦИИ И ДИФФЕРЕНЦИАЦИИ КЛЕТОК И ТКАНЕЙ
Обратимся вновь к тому, что совершается в ходе нормального развития, — к явлениям дифференциации и к анализу процессов, названных Шпеманом индукцией, и разберем, что, собственно, является взаимозависимостью частей в развитии.
В 50-х и 60-х годах XX в. в связи с возрастающим взаимовлиянием биологии, физики и химии и с использованием новых методик (ауторадиография, электронная микроскопия и др.) снова повысился интерес к химии индукторов, хотя содержание этого понятия резко изменилось. Во-первых, считается неосновательным искать какое-либо одно формообразовательное вещество, вызывающее индукцию. Во-вторых, все меньшее число исследователей уподобляет явления индукции, наблюдаемые в ходе нормального развития зародышей, феномену мертвых организаторов. В-третьих, вместо гипотезы Шпемана об индукционных влияниях организатора на «индифферентный» клеточный материал утвердилась мысль о взаимозависимости частей в развитии зародышей.
В течение длительного периода, последовавшего после увлечения и разочарования химическими исследованиями (40-е и 50-е годы), некоторые эмбриологи продолжали изучать природу Индукторов. К ним относятся прежде всего С. Тойвонен и Т. Ямада, которые вместе с другими учеными создали качественную гипотезу индукторов. Тойвонен испытал действие сотен тканей разных животных на их способность к индукции осевых зачатков у амфибий. В 1938 г. он обнаружил, что некоторые индукторы обладают качественно различным действием: ткань печени морских свинок индуцирует почти исключительно передний мозг и его производные (сначала передний мозг, затем глаза и балансеры). Костный мозг морской свинки индуцируют туловищные и хвостовые структуры (миотомы, хорду, почечные канальцы и т. д.).
Ф. Леман в 1950 г. предложил гипотезу, принятую Тойвоне- ном, Ямадой ц другими исследователями. Согласно ей, первичная индукция (сущность действия организатора — дорсальной губы бластопора) может быть вызвана всего двумя агентами, образующими два взаимоперекрывающихся градиента. Одно вещество индуцирует исключительно переднеголовные (архенцефалические) структуры, а другое—туловищно-хвостовые (дейтеренцефаличес- кие) структуры. Если много архенцефалического агента и мало дей- теренцефалического, то индуцируется передний мозг; если первого агента относительно мало, а второго больше, то возникает
заднеголовная область; если мало переднеголовного агента и много туловищно-хвостового, то индуцируются туловищно-хвостовые отделы. Все это и имеет место, согласно гипотезе, в нормальном развитии амфибий; надо представить себе наличие определенных индуцирующих веществ в соответствующих количественных комбинациях в разных участках зародыша.
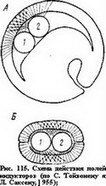



 С. Тойвонен и Л. Саксен в 1955 г. проверили эту гипотезу экспериментально, поставив опыты на зародышах амфибий с одновременной и раздельной индукцией — печенью и костным мозгом. Ткани печени и костного мозга или то и другое вместе пересаживали в гаструлу тритона или завертывали в. эктодерму гаструлы тритона (метод «сэндвичей»). Как видно из схемы на рис. 115, ткань печени индуцирует преимущественно образование переднего мозга и его производных и изредка также заднего мозга со слуховыми пузырьками; индукция мезодермы не наблюдается. Костный мозг индуцирует главным образом мезодер- мальные структуры; головные структуры не образуются. В опытах с одновременным действием печени и костного мозга возникли структуры всех уровней тела нормальной личинки. В ряде случаев на брюшной стороне хозяина развивались полные вторичные зародыши, подобные тем, которые возникают нередко и при трансплантациях верхней губы бластопора или при действии мертвых организаторов.
С. Тойвонен и Л. Саксен в 1955 г. проверили эту гипотезу экспериментально, поставив опыты на зародышах амфибий с одновременной и раздельной индукцией — печенью и костным мозгом. Ткани печени и костного мозга или то и другое вместе пересаживали в гаструлу тритона или завертывали в. эктодерму гаструлы тритона (метод «сэндвичей»). Как видно из схемы на рис. 115, ткань печени индуцирует преимущественно образование переднего мозга и его производных и изредка также заднего мозга со слуховыми пузырьками; индукция мезодермы не наблюдается. Костный мозг индуцирует главным образом мезодер- мальные структуры; головные структуры не образуются. В опытах с одновременным действием печени и костного мозга возникли структуры всех уровней тела нормальной личинки. В ряде случаев на брюшной стороне хозяина развивались полные вторичные зародыши, подобные тем, которые возникают нередко и при трансплантациях верхней губы бластопора или при действии мертвых организаторов.Тойвонен предполагает, что каждый из двух индукторов образует свое активное поле, при одновременном их действии возникнет комбинированное поле (рис. 115).
На основании подобных опытов Тойвонен представляет себе существование такого индукционного механизма у зародышей.
Предположения и данные разных исследователей о химической природе N- и М-агентов очень противоречивы. Предполагают, что М- и N-агенты — это белковые соединения. X. Тидеман (1968) извлекал из гомогенатов девятидневных куриных зародышей индуцирующий мезодермальный фактор белковой природы (относительная молекулярная масса 25000—30 000). Он инактивируется тиогликолевой кислотой или меркаптоэтанолом, значит, содержит сульфидный мостик цистина. При имплантации этого вещества в гаструлу амфибии происходило во многих случаях формирование мезодермальных структур. Трудно, однако, говорить об этом веществе как о специальном «мезодермальном факторе», так как в опытах кроме формирования мышщ- хорды и почечных трубочек образовывались и энтодермальные ткани. Считают, что индукция архенцефалических структур может быть получена рибонуклеопротеинами. Однако чистые протеины из костного мозга и куриного эмбриона действуют как каудально-мезо- дермальные индукторы.
Анимализации можно добиться уменьшением хирургическим путем вегетативного материала или обработкой перед оплодотворением некоторыми химическими веществами, такими, как KCN, или обработкой в период дробления иодобензойной кислотой или 2,5-тиометилцитозином.
Вегетализация, т. е. более интенсивное развитие энтодермаль- ных зачатков, может быть вызвана обработкой дробящихся яиц литием или хлорамфениколом. К 70-м годам химия «индукторов» оказалась столь же неясной, как и в период односторонних химических увлечений эмбриологов в 30-е годы.
Подробно изложенная интересная гипотеза Тойвонена не дает, к сожалению, ничего принципиально нового по сравнению со старыми односторонне химическими схемами сущности индукторов и организаторов, только вместо одного вещества думают о двух или нескольких. Должны быть приняты во внимание следующие очевидные недостатки гипотезы Тойвонена, на которые отчасти указывает и сам автор. Во-первых, эта гипотеза говорит только
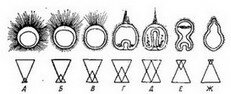
Рис. 117. Вариации проявления тенденций «анимализации» и «вегетализации» развивающихся яиц морских ежей (по Т. Густафсону, 1965):
А—Г — различные случаи «анимализированных» личинок; Д — нормальная гаструла; Е— Ж — различные случаи «вегеталнзированных» личинок. В случае «Ж» эктодерма редуцирована до маленькой «почки». Анимализация (редукция вегетативного материала) производилась или обработкой химическими веществами перед оплодотворением (например, SCN), или обработкой в ходе дробления яйца иодобензойиой кислотой или 2,5-тиометилцнтозином. Вегеталнзацня производилась обработкой солями лития или хлорамфениколом по ходу дробления яиц
об «индукторах» и совершенно не касается главного вопроса — о «реагирующих» системах. Во-вторых, экспериментальное обоснование ее дано на основании действия каких-то веществ тканей млекопитающих, а делается попытка объяснений явлений нормального развития зародышей амфибий. Требуется доказать, что выделенные Тойвоненом, Ямада и другими «индуцирующие вещества» действительно присутствуют в нормальной гаструле. Если они присутствуют, то каково их местоположение? Поэтому к гипотезе Тойвонена и сходным с ней относятся все те критические замечания, которые были уже сделаны о шпемановской школе. С данными Тойвонена перекликаются результаты опытов по экзо- гаструляции (рис. 117).
Источник: Токин Б. П.. Общая эмбриология: Учеб, для биол. спец, ун-тов.—4-е изд., перераб. и доп. 1987
Еще по теме ХИМИЧЕСКИЙ АСПЕКТ ИЗУЧЕНИЯ ДЕТЕРМИНАЦИИ И ДИФФЕРЕНЦИАЦИИ КЛЕТОК И ТКАНЕЙ:
- Часть II. ИЗУЧЕНИЕ СТРОЕНИЯ И ЖИЗНЕДЕЯТЕЛЬНОСТИ КЛЕТКИ И ТКАНЕЙ, НАСЛЕДСТВЕННОСТИ И ИНДИВИДУАЛЬНОГО РАЗВИТИЯ ОРГАНИЗМОВ
- Глава I ОБ ЭКОЛОГИЧЕСКИХ АСПЕКТАХ ИЗУЧЕНИЯ ПОВЕДЕНИЯ
- Физико-химическое изучение клетки
- Применение органического анализа для изучения химических процессов в организме
- 8.3. ЦЕЛОСТНОСТЬ ОНТОГЕНЕЗА 8.3.1. Детерминация
- Успехи в изучении коферментов. Доказательство химической общности коферментов и витаминов
- 7.5.3. Образование органов и тканей
- Резорбция из тканей
- Теория дивергентной эволюции тканей
- 3.5.2. Физико-химическая организация хромосом эукариотической клетки 3.5.2.1. Химический состав хромосом
- 15.4. ВНУТРИВИДОВАЯ ДИФФЕРЕНЦИАЦИЯ ЧЕЛОВЕЧЕСТВА
- 8.2.5. Дифференцировка клеток
- Функциональная дифференциация организма
- 8.2.3. Сортировка клеток
- 8.2.2. Миграция клеток
- 8.2. МЕХАНИЗМЫ ОНТОГЕНЕЗА 8.2.1. Деление клеток
- Дифференциация Человека разумного на расы
- 8.2.4. Гибель клеток