Функциональный анализ роста
Функциональный анализ роста исходит из нескольких основных параметров, которые преимущественно касаются того, как ассимиляты распределяются в растении по различным органам и как они в них размещены.
Окружающая среда может влиять на процессы распределения таким образом, что ассимиляты предпочтительно инвестируются там, где имеется их недостаток: при сухости почвы — в корни, при недостатке света — в листья и т.д. Это проявляется в сухой массе органов по отношению к обшей сухой массе растения (рис. 13.34). В английской номенклатуре для этого применяются следующие понятия:- LMF — leaf mass fraction (листовая часть);
- SMF — stern mass fraction (масса всех побегов);
- RMF — root mass fraction (корневая часть).
Аналогично в качестве фракций биомассы могут быть обозначены репродуктивные органы, запасающие органы и др. В последнее время умножаются доказательства того, что эти модели распределения биомассы не всегда соответствуют впервые описанному Р. Броуером «функциональному равновесию» и распределение сухого вещества очень сильно реагирует на факторы окружающей среды. Как правило, к этой проверенной модели добавляются еще физиологические и морфологические коррекции (например, регулирование насыщения азотом и продуктивность фотосинтеза в листьях, кутикула и размеры листьев).
Fla уровне листа и корня нас интересует «стоимость» биомассы, необходимая для создания функционального единства. Так как функция листьев заключается прежде всего в абсорбции света, она описывается лучше через листовые поверхности. У корней это интенсивность пронизывания почвы, следовательно, длина тонких корней, соответствующая единице биомассы. Именно поверхность корней вступает в контакт с почвой, правда, качество (активность) этой поверхности меняется с возрастом корней. В отношении к корневой поверхности нельзя занижать оценку роли очень активных абсорбирующих тонких корней и переоценивать роль уже не абсорбирующих (преимущественно проводящих) толстых старых корней.
Поэтому независимо от толщины часто используют длины корней как функциональную меру. Соответствующие параметры обозначаются как:- SLA (specific leaf area — специфическая листовая поверхность, квадратный метр поверхности листьев на грамм их сухого веса, или для наглядности— дм2-г' или м2 - кг"1);
- SRL (specific root length — специфическая длина корней; метр длины корней на грамм их сухого веса).
Вместо SLA часто употребляют обратное этому показателю значение LMA (leaf mass per area). Из сочетания долей массы органов и их специфической «стоимости» получают два важных уравнения функционального анализа роста:
LAR = LMF SLA
(leaf area ratio — общая поверхность всех листьев по отношению к общей биомассе всего растения, м2-г"’);
RLR = RMF SRL
Деревья Кустарники Многолетники ^уліЗІГ'0 Однолетники
80-98%
В фотосинтезирующих органах находятся очень различные доли от общей биомассы. Здесь приведена leaf mass fraction (LMF), как отношение листовой фракции биомассы к общей биомассе (% сухой массы, затемненная плоскость) взрослых растений различных морфотипов, т.е. разных жизненных стратегий
Между тем многочисленные исследования подтверждают, что LAR представляет собой важнейший детерминант роста (рис. 13.35), причем решающими переменными для значения LAR могут быть как LMF, так и SLA.
LAR — прежде всего статичная величина. Необходима еще оценка специфической продуктивности фиксации углерода листовой поверхностью. Она будет обозначена как ULR (unit leaf rate) и описывается приростом сухого веса всего растения по отношению кім2 листовой поверхности в день (г м 2-день-1). Отсюда вытекает и относительная степень роста RGR одного растения:
RGR = UFR - FAR.
ULR представляет собой интегральную меру для ассимиляционной продуктивности листьев по отношению к достигнутому с ее помощью нетто-приросту растения и отражает ассимиляцию углерода более реалистично, чем кратковременное измерение листового фотосинтеза.
Недостаток ULR состоит в том, что она определяется только деструктивно (прирост массы за относительно короткие, например, недельные интервалы) и поэтому ее можно установить только для травянистых или во всяком случае мелких растений. Для деревьев в грубом приближении могут быть сопоставлены с LAI прирост древесины и продукция опа- да. Так как при росте большую роль играет эффект «сложного процента» и ход кривой роста во времени редко бывает линейным, делить годовой прирост растения на длительность вегетационного периода и считать результат этого деления за RGR некорректно. Употребляющаяся в сельском хозяйстве мера продуктивности роста растения за более длительные периоды (несколько недель, месяцы) — расчетный синоним ULR, интенсивность нетто- ассимиляции (NAR — net assimilation rate) — обозначает прирост биомассы на усредненную листовую поверхность в течение одного, как правило, более долгого периода наблюдений. Однако отношение к быстроменяющейся за этот длительный промежуток времени листовой поверхности проблематично.RGR варьируется в зависимости от вида двумя порядками величин. У деревьев LMF и RMF более-менее условно, так как прирост благодаря включению массивной неактивной ядровой области древесины будет очень мал. По отношению к активной проводящей части древесины LMF и RMF будут иметь почти те же значения, что и у многолетних травянистых растений. Из-за ежедневной «уплаты процентов» на прирост листьев RGR может очень быстро достичь крайне высоких значений. 20 % прироста в день у молодых травянистых растений не редкость. Многие анализы между тем подтверждают, что быстро и медленно растущие растения различаются преимущественно в
0,4 -
Рис. 13.35. Регулирование роста путем ассимиляции и инвестиций.
Относительная интенсивность роста варьирует независимо от интенсивности ассимиляции С02 суммой всех листьев (ULR — unit leaf rate), но коррелирует линейным образом с отношением листовой поверхности к общей биомассе (LAR — leaf area ratio).
Зависимость от LAR принадлежит преимущественно SLA-компоненту (поверхность листьев на их сухую массу), а не отношению массы листьев к общей биомассе (LMF). Данные по 51 разному травянистому растению при оптимальных условиях роста сведены H.Poorter и A. Van der WerfТаблица 13.2. Параметры* функционального анализа роста
|
Тип растительности |
LMF |
SMF |
RMF |
SLA |
SPL |
LAR |
RGR |
|
Травянистые растения |
0,25 |
0,45 |
0,30 |
25 |
50 |
6 |
0,15 |
|
Сезонно-зеленые деревья |
0,02 |
0,85 |
0,13 |
12 |
— |
0,24 |
0,02 |
|
Вечнозеленые хвойные деревья |
0,04 |
0,83 |
0,13 |
3 |
— |
0,12 |
0,02 |
* Грубые контрольные цифры для уже хорошо развитых, но еще не сенильных особей в природе; у всходов, ювенильных и сенильных особей численные значения могут сильно отклоняться. LMF, SMF, RMF даны в г г1, LAR— в м2-кг' и RGR— в г-г1-день'1 (все по отношению к сухому весу всего растения), SLA— в м2- кг1, SPL— в м-г 1 в единицах веса сухой ткани (листьев или корней).
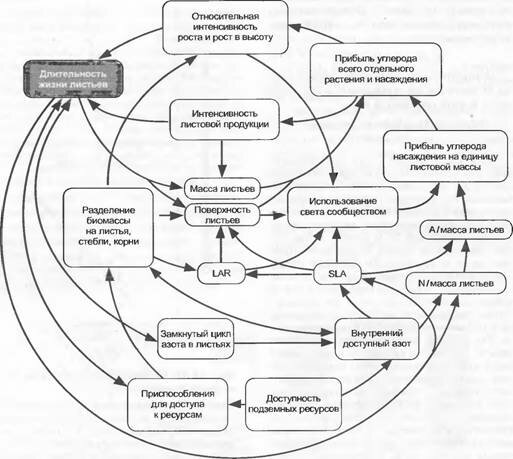
Рис. 13.36. Модель функциональной связи распределения и ассимиляции Си N с длительностью жизни листьев (по P. Reich).
Приняты во внимание только внутренние регуляторы роста растения и доступности питательных веществ без включения таких внешних факторов, как климат и влажность почвы, а также процессы развития и биотические взаимодействия (А — продукция фотосинтеза, LAR — leaf area ratio, SLA — specific leaf area)
инвестициях биомассы относительно листовой поверхности и суммарной длины корней.
Быстро растущие растения в противоположность медленно растущим имеют большие значения SLA (см. табл. 13.1 в разделе 13.6.2) и SRL. Типичные значения этих параметров приведены в табл. 13.2.Рис. 13.36 схематически иллюстрирует согласованность важнейших детерминантов функционального анализа роста и при этом показывает (дугообразными стрелками) отношения отдельного листа к прибыли углерода в монокультуре. Эта функциональная схема была бы еще сложнее, если бы попытались включить многогранные (в значительной степени неизвестные) взаимодействия с симбионтами, фитофагами, патогенными организмами и деструента- ми и понять механизм процесса развития растения (например, цветения), принимая во внимание эти факторы. Комплексность еще более повышается, если и на уровне сообщества также принять в расчет надземные и подземные взаимодействия между различными видами растений и возрастными группами. Рост растительного сообщества поэтому нельзя предсказать (смоделировать) на механистической основе, исходящей из функций одного листа. Для этого отсутствуют данные по бесчисленным параметрам. Такие параметры совершенно непонятны, поскольку они представляют собой переменные, зависящие от других переменных. Часто используемые представления о механической системе из шестеренок и приводных ремней ведут к ошибкам, так как в реальном мире растения «колеса при работающем моторе, его величина и форма, а также число зубцов варьируют, приводные ремни растягиваются в направлении применяемых усилий», и кроме того, рабочая программа (развитие) быстро меняется. Каждый прогноз роста растения имеет поэтому статистическую природу и основывается на тенденциях (вероятностях), которые в такой же степени выводятся из статистических наблюдений в прошлом. В растениеводстве это давно известно. Показанная здесь комплексность проясняет, почему нельзя ожидать того, что путем генетического вмешательства в какой-нибудь частный процесс (например, в процессы, происходящие на мембранах хлоропластов) можно будет в естественных условиях получить прогнозируемые результаты роста.
Еще по теме Функциональный анализ роста:
- Модели роста популяций.
- Экология роста
- 1. Стимуляторы роста растений.
- Особенности роста и развития растений кукурузы
- РЕГУЛЯТОРЫ РОСТА РАСТЕНИЙМИКРОБНОГО ПРОИСХОЖДЕНИЯ
- Глава 10 Ранние этапы роста семь
- СПОСОБЫ ОПТИМИЗАЦИИ МИКРОКЛИМАТА И СТИМУЛЯЦИИ РЕЗИСТЕНТНОСТИ, РОСТА, РАЗВИТИЯ И ПРОДУКТИВНОСТИ животных
- Принципы и типы функциональной эволюции
- ПИЩЕВАРИТЕЛЬНЫЙ ТРАКТ (морфологический и функциональный аспекты)
- Злобин Ю. А.. Популяционная экология растений; современное состояние, точки роста ; монография. - Сумы ; Университетская книга. - 263 [2Jc., 2009
- Анализ по С.П. Мартынову