Передача информации о корме
Прос/ейший способ передать другим особям сведения о найденном источнике корма состоит в том, чтобы, вернувшись в гнездо, совершить определенные демонстративные действия.
Одновременное выделение секрета из мандибулярных желез может активизировать потенциальных фуражиров и снабдить их запаховыми указаниями о типе корма и его местонахождении. Если гнездо имеет несколько входов, то вход, к которому вернулся фуражир, может указывать, в каком секторе лучше всего вести поиски. Дальнейшими указателями могут служить пахучие метки-секреты мандибулярных желез или экскременты,- оставляемые где-нибудь снаружи, так что прямая, соединяющая выход из гнезда с такой меткой, указывает направление, в котором надо следовать. Стимулирующие химические вещества содержатся и в метках, оставляемых с помощью других желез. Для большей надежности особь, обнаружившая пищу, может собрать группу рабочих и вывести их из гнезда, используя свою зрительную топографическую память и метки, чтобы вернуться к найденному источнику корма и помочь другим не терять связи между собой.Наиболее совершенная из всех выработанных в процессе эволюции систем - это указание направления при помощи пробежки, выполняемой в гнезде; при этом передается даже информация о расстоянии до источника корма: частота вибраций, совершаемых при пробежке, коррелирует с временем, необходимым для того, чтобы добраться до этого источника.
У большинства ос сообщение о найденном корме сводится к тому, что оса проявляет некоторое оживление, вернувшись в гнездо; возможно, что при этом передаются какие-то указания при помощи запахов, как у Vespula (Maschwitz et al„ 1974; цит. no Jeanne, 1980), но у Vespa помощники могут следовать за лидером, возвращающимся к корму. Пчелы Meliponinae оставляют простые метки, например представители рода Trigona на пути от источника корма к гнезду наносят на растения пахучие метки из выделений мандибулярных желез.
Они делают это медленно и методично, а на некотором расстоянии от гнезда метки кончаются, причем расстояние это видоспецифично: так, у T.postica оно равно 8 м, а интервалы между метками составляют по 1 м (Lindauer, Кегг, 1958, 1960). У этого вида, как и у всех других, секрет мандибулярных желез представляет собой смесь летучих соединений, в которой преобладает бензойный альдегид, но содержатся также тридекан-2-он и пентадекан-2-он.У Т. subterranea секрет состоит из альдегидов монотерпенового ряда (нераля и гераниаля - двух стереоизомеров цитраля), а у Scap to trigona -из карбонильных соединений (Blum, Brand, 1972). Вернувшись в гнездо, эти насекомые возбуждают и активизируют других зигзагообразными пробежками и жужжанием, которые видоспецифичны, но не несут конкретной информации. Затем пчела-информатор летит к источнику пищи, сопровождаемая завербованными ею сборщицами. Таким образом, мобилизация сборщиц у Trigona включает три элемента: оставление следа, зигзагообразные пробежки и следование за лидером.
АІуравьи Ponerinae, подобно мелипонинам, чтобы привлечь помощников, используют мандибулярные железы. Odontomachus troglodytes, обладающий сильно развитым жалом, нападает на жертву, равную по размерам ему самому; кусая и жаля других особей своего вида, он заставляет их оказывать ему помощь в перетаскивании жертвы, а выделяемая им смесь алкилпиразинов активизирует других муравьев, привлекает их и побуждает к нападению (Longhurst et al., 1978). Когда муравей Bothroponera soror находит крупную жертву, его мандибулярные железы выделяют феромон, привлекающий других рабочих, находящихся в радиусе 20 см. В состав этого феромона входит не менее 21 соединения, однако все те реакции, которые он вызывает,- вытягивание антенн вперед, приближение, смыкание челюстей и вонзание жала-можно вызвать при помощи смеси ундекан-2-ола, 2-метил-6-метилсалицилата и ундекан-2-она в соотношении 40 :10 :1. Муравьи этого вида также привлекают помощников к тому месту, откуда нужно перетащить очень крупную добычу (Longhurst et al., 1980), но у других видов рабочая особь вербует себе помощников, возвращаясь в гнездо и приглашая других муравьев следовать за собой; она выбегает наружу, а другой муравей бежит за ней, прикасаясь к ее брюшку и ногам.
Такому „тандему" способствует пигидиальная железа, расположенная между сегментами в дорсальной части брюшка (Holldobler, Traniello, 1980а;Maschwitz et al., 1974); эта железа может использоваться для создания пахучего следа: бегущий муравей загибает кончик брюшка вниз и трется им о землю, оставляя за собой след. У некоторых муравьев такое поведение дает возможность мобилизовать сразу нескольких рабочих, которые следуют по „одноколейному" пути, проложенному разведчиками (Fletcher, 1973;'Holldobler, Traniello, 1980b).
У муравья Megaponera foetens (Ponerinae), облигатно питающегося термитами, кривая распределения рабочих особей по размерам трехвершинная (van Boven,
- ; этот муравей ориентируется по кронам деревьев (Holldobler, 1980). Обнаружив свежие следы деятельности термитов в почве, муравей-разведчик прокладывает мобилизационный след к своему гнезду. Спустя примерно 5 мин он снова выходит наружу в сопровождении нескольких крупных и множества мелких рабочих; если на этом этапе удалить разведчика, то возникнет хаос, несмотря на то что имеется проложенный путь (рис. 4.1; Longhurst, Howse, 1978). Когда муравьи доходят до зоны термитов, крупные рабочие снимают слои почвы, а мелкие проникают в почву и вытаскивают из нее термитов, складывая их в кучки снаружи, пока операция не закончится. По-видимому, нападение координируется и сосредоточивается в одном месте благодаря выделению мандибулярными железами феромона, состоящего главным образом из диметилдисульфида и диметилтрисульфида. Спустя примерно 10 мин муравьи выстраиваются в колонну и возвращаются сомкнутыми рядами, по 3-5 муравьев в ряду; крупные рабочие идут в авангарде и готовы отбить нападение грабителей. На обратном пути муравьи движутся со скоростью 4,6 см/с-быстрее, чем они шли от гнезда (3,8 см/с) (Fletcher, 1973;
ние может контролироваться разведчиками. Объектом охоты служат термиты Macrotermes. (Longhurst, House, 1979а.)
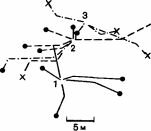
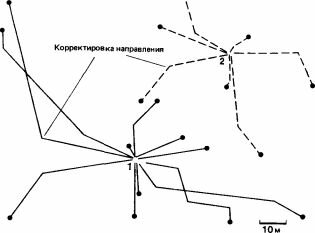
Рис. 4.1. Фуражировочные следы Megaponera foetens. А. Семья, у которой следы начинаются от двух входов гнезда. Б. Семья, у которой следы начинаются от трех входов. Направле
Longhurst, Howse, 1978, 1979a; Longhurst et al., 1979). Муравьи рода Megaponera, подобно многим другим Ponerinae, производят довольно хорошо слышное стрекотание (Markl, 1973), главным образом если их потревожить или поймать, однако неизвестно, помогает ли это им при фуражировке. В Кении наблюдается три пика охотничьей активности в день; в среднем один муравейник поедал ежедневно 653 термита, главным образом Macrotermes subhyalinus (Lepage, 1981а).
Подсемейство Myrmicinae отличается большим разнообразием, чем Ponerinae. Представители рода Myrmica обитают в травянистых и кустарниковых зонах умеренного пояса и питаются падью, живыми мелкими животными и их трупами; при фуражировке и других передвижениях они пользуются зрением, ориентируясь по контуру горизонта, отдаленным объектам, поляризации света неба или положению солнца. Его реакции на пищу варьируют: обнаружив воду или мелкую добычу, муравей-разведчик прокладывает с помощью жала ядовитый след и возвращается в гнездо нагруженным [Kugler (1979а) описывает эволюцию жала в „писчее перо" у Myrmicinae], В гнезде он ведет себя спокойно, побуждая лишь нескольких рабочих выйти и направиться по оставленному им следу. Если же обнаружена крупная жертва или много мелких, то разведчик сначала оценивает ситуацию, а затем возвращается в гнездо без груза; как и в первом случае, он оставляет метки, выделяя яд из жала, но добавляет к этому секрет дюфуровой железы, особенно вблизи гнезда и вокруг найденной пищи. Войдя в гнездо, он начинает энергично и быстро бегать, бросаясь из стороны в сторону, так что десятки рабочих устремляются по следу в сопровождении разведчика или без него (Cammaerts, 1978). Яд представляет собой в основном раствор белков и аминов, но содержит также слабо полярное низкомолекулярное летучее соединение-следовой феромон (Cammaerts-Tricot et al., 1977).
В дюфуровой железе синтезируются линейные углеводороды с цепью из 13-19 атомов углерода, три фарнезина (терпеноиды) и четыре низкомолекулярных вещества, содержащих кислород: ацетальдегид, этанол, ацетон и бутанон в соотношении 35 :3 :40 :25 (Morgan et al., 1979). Эта смесь постепенно испаряется, начиная с низкомолекулярных спиртов, альдегидов и кетонов, которые возбуждают рабочих и указывают им направление; затем испаряются терпеноиды и, наконец, углеводороды. Такая последовательность дает рабочим возможность оценить свежесть следа. Так обстоит дело у Myrmica rubra и М. scabrinodis, причем муравьи этих двух видов могут следовать по дорогам, проложенным особями как своего, так и другого вида. Однако, хотя дюфурова железа у М. scabrinodis секретирует те же летучие вещества, что и у М. rubra, в секрете этого последнего вида содержатся иные, менее летучие соединения, используемые для того, чтобы метить территорию (Cammaerts et al., 1977, 1978); при нанесении меток муравей медленно передвигается по территории, оставляя капельки секрета. Мандибулярные железы видов Myrmica вырабатывают также много химических веществ, которые выделяются при встрече с жертвой и привлекают на помощь находящихся поблизости сотоварищей по гнезду (Morgan et al., 1978; Parry, Morgan, 1979). Например, у M. scabrinodis секрет этих желез содержит 49% октан-3-она, 18% октан-3-ола, 12% декан-3-она и по меньшей мере 16 близких к ним соединений-от этанола до ундекан-3-она. У М. rubra имеется сходный набор соединений, но в иных соотношениях (Cammaerts et al., 1981а).
У Monomorium venustum рабочие особи, для того чтобы заставить других рабочих выйти из гнезда, устраивают довольно сложную демонстрацию, состоящую из особых движений-виляний из стороны в сторону, толчков головой, ударов усиками и судорожных пробежек. По мере иссякания источника корма демонстрация упрощается: сначала отпадает виляние, затем удары усиками и, наконец, пробежки (Szlep, Jacobi, 1967). Crematogaster, перешедший к жизни на деревьях примерно в середине третичного периода (Brown, 1973), в настоящее время использует для нанесения меток секрет особых тибиальных желез, находящихся на задних ногах, которыми он быстро и энергично барабанит по субстрату, с тем чтобы кончики лапок распространяли феромоны (Fletcher, Brand, 1968;
Leuthold, 1968). Один из видов Novomessor может привлекать помощников с расстояния до 2 м, поднимая брюшко и выделяя из кончика жала летучее вещество; он также выводит наружу своих сотоварищей по гнезду, прокладывая ядовитый след, но не использует для оставления следов пигидиальную железу. Стридуляция создает звуки, распространяющиеся через субстрат и усиливающие реакцию на феромоны (Holldobler et al., 1978; Markl, Holldobler, 1978).
Аналогичный ряд последовательных этапов в процессе мобилизации существует у Formicinae. Wallis (1964) не смог обнаружить никаких признаков мобилизации помощников у Formica fusca, однако возможно, что эти муравьи прокладывают мобилизационный след при помощи материала, выделяемого задней кишкой, и возбуждают потенциальных фуражиров, производя виляющие движения (Moglich, Holldobler, 1975). Camponotus pennsylvanicus ведет себя подобно F. fusca, с той разницей, что он добавляет к следу еще муравьиную кислоту (Traniello, 1977). С. socius уделяет больше внимания вербовке помощников; разведчик прокладывает след из выделений задней кишки (возможно, это экскременты с добавлением секрета ректальных желез), ведущий прямо к гнезду, устраивает в гнезде демонстрацию с вилянием брюшком и предложением пищи, собирает несколько десятков помощников, а затем ведет их за собой по следу; муравьиная кислота, выделяемая ядовитой железой разведчика, активизирует следующих за ним особей (Holldobler, 1971).
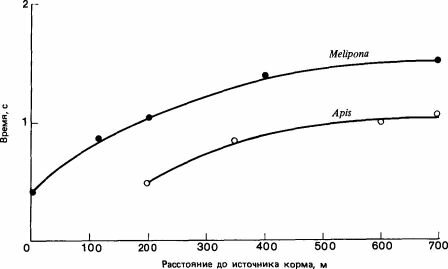
Рис. 4.2. Соотношение между расстоянием до источника пищи и длительностью звукового сигнала у Apis mellifera и Melipona quadrifascia- ta. (Esch et al., 1965.)
У С. sericeus разведчик медленно выходит наружу, а за ним следует лишь одна особь, прикасающаяся к нему; в случае утраты контакта ведущий останавливается, чтобы восстановить связь, используя след в качестве путеводной нити (Holldobler et al., 1974; Maschwitz, 1975a).
Melipona - один из родов безжалых пчел-ведет группы рабочих особей к корму после того, как поднимет других пчел жужжанием и пробежками; разведчица движется зигзагами в нужном направлении, а за ней летят другие пчелы. Они пролетают таким образом всего 30-50 м (М. quadrifasciata) или 10-20 м (М. seminigra). Хотя теоретически этого достаточно, чтобы установить направление, требуется повторить такой путь много раз, прежде чем мобилизованные рабочие-помощницы начнут искать корм дальше того места, до которого их доводят (Holldobler, 1977). Длительность жужжания коррелирует с расстоянием до корма (рис. 4.2), поэтому возможно, что сведения о расстоянии передаются при помощи звуков, хотя это еще не подтверждено (Esch, 1967; Esch et al., 1965).
Apis mellifera тоже издает жужжание, продолжительность которого пропорциональна расстоянию до источника корма и которое передается восприимчивым пчелам в виде вибрации сота (Wenner, 1962). Этим жужжанием пчелы сопровождают свой знаменитый «виляющий танец» (в основном это пробежка по прямой, во время которой брюшко движется из стороны в сторону). С расстоянием до корма коррелирует также то, на какой глубине в улье исполняется танец, его продолжительность, длина прямой пробежки и число совершаемых при этом виляний. Таким образом, танец содержит избыточную информацию, что характерно для языков вообще, и трудно решить, какой элемент (если таковой существует) играет наиболее важную роль[III].
Apis Jlorea часто танцует на площадке поверх своих сотов и способна указать, в каком направлении находится пища; разумеется, расстояние при этом уменьшено (Free, Williams, 1979).
A. dorsata танцует на своих вертикальных сотах, но преобразует указание «к солнцу» в указание «вверх»: направление указывается как угол, который оно образует с направлением «вверх», а не «к солнцу», т.е. так же, как у A. mellifera. Такое преобразование свойственно не только пчелам; муравьи, бежавшие по горизонтальной пластинке под определенным углом к солнцу, побегут под тем же углом к направлению вверх, если поставить пластинку вертикально (Markl, 1964).
При этой системе не учитывается высота источника корма над землей.
Однако, обнаружив источник корма, расположенный на некоторой высоте или на земле вблизи улья, пчела выполняет «круговой танец», сообщая своим подругам, что следует искать пищу поблизости. Медоносные пчелы разных подвидов исполняют круговой танец при разных расстояниях до источника корма: A. mellifera ligustica- в том случае, если корм находится в радиусе 100 м, А. т. carnica- в радиусе 85 м, А. т. fasciata- 12 м, A. indica - всего лишь 2 м (Holldobler, 1977). При больших расстояниях наблюдается постепенный переход к виляющему танцу. Медоносные пчелы обладают прекрасной памятью, хорошо развитым чувством времени, хорошим обонянием, цветовым зрением и способностью легко осваиваться с процедурой эксперимента, а также с присутствием и действиями экспериментатора (Wells, 1973; Wenner, 1974). Помимо внешнего вида и запаха экспериментатора пчеле приходится привыкать к запахам, возникающим, когда люди топчут растения, скашивают траву или метят пчел красками, а также осваиваться с различиями в естественной растительности и ландшафте. Если приучить пчел к раствору сахарозы с определенным запахом, а затем дать раствору высохнуть, то достаточно вдуть немного душистого воздуха в улей, чтобы пчелы вылетели и устремились к знакомому блюдцу или к любому другому блюдцу, издающему тот же аромат. Пчел привлекают также другие пчелы-как своим видом, так и запахом, который от них исходит; возможно, это тот же запах, что в улье и у входа в улей (Ferguson, Free, 1979). Таким образом, существует много источников ошибок при проведении экспериментов, и необходим тщательный контроль.
Чтобы избежать всего этого, Gould (1976) использовал то обстоятельство, что пчелы ориентируют свой танец относительно внутреннего источника света в улье таким же образом, как если бы это было солнце. Заклеивая разведчицам глаза, он смог понизить их чувствительность к источнику света, помещенному в улье, так что они не обращали на него внимания и исполняли танец как обычно, направляясь вверх по сотам; однако мобилизованные ими новички, способные видеть свет, так как их глаза не были заклеены, воспринимали указания о направлении неверно.
Реакция на запах не оказала бы такого влияния.Паратион сокращает длину прямого пробега при виляющем танце, который при этом неверно указывает более близкое расстояние, чем на самом деле; пчелы следуют этому указанию и направляются к более близкому месту, чем надо, игнорируя сигналы, поступающие от местных запахов (Gould, 1976). В настоящее время нет сомнений в том, что медоносные пчелы общаются друг с другом при помощи символов, однако все еще остается неясным, в какой мере они их используют. Очень часто танцы, сообщающие об источниках корма, не привлекают внимания; на них реагируют только молодые, неопытные сборщицы, причем им надо наблюдать эти танцы по многу раз, чтобы понять заключенное в них сообщение. У более старых пчел, у которых уже выработались определенные привычки, по-видимому, имеется пристрастие к тем или иным цветкам, раскрывающимся в разное время дня; поэтому они сидят в каком-нибудь укромном уголке улья, пока одна из вернувшихся разведчиц не сообщит, что «их цветок» готов; сигналом здесь может быть только запах. Во всяком случае, пчелы
примерно знают, в какое время дня раскрывается тот или иной цветок, и ждут подтверждения от возвращающихся разведчиц.
Небольшие ошибки в танцах пчел, указывающих направление, можно объяснить наличием в передней части брюшка магнитных гранул, чувствительных к магнитному полю Земли. Эти ошибки можно устранить наложением магнитного поля противоположного знака. Кроме того, Tomlinson et al. (1981) показали, что если около пчелы, исполняющей обычный танец, поместить магнит, то продолжительность танца сокращается с 34 до 19 с; это подтверждает, что пчелы чувствительны к магнитному полю, и показывает, что очень неоднородное поле тоже может повлиять на танец. Наконец, у Meliponini наблюдается даже элемент мобилизации путем привода; после того как две группы меченых пчел получали корм на кормушках, расположенных на разной высоте и закрытых ширмой, некоторые из этих пчел возвращались из гнезда, сопровождаемые новыми (немечеными) особями, которым они, очевидно, служили проводниками (Wellington, Cmiralova, 1979). Описаны и другие эксперименты, подтверждающие это представление (Friesen, 1973).
Apis mellifera издает запах, исходящий от желез Насонова (возможно, гомологичных пигидиальным железам муравьев); этот запах распространяется в воздухе и, по-видимому, используется в тех случаях, когда природных запахов мало, например при движении роя и образовании роевого клуба (Free, 1968). Важную роль играют также менее летучие вещества, распространяемые лапками пчел, когда они ходят около корма, поедают его или же входят в улей (Ferguson, Free, 1979). Эти вещества специфичны для каждой семьи и, по-видимому, выделяются межсегментными железами, расположенными на дорсальной стороне брюшка, а не железами Насонова; оно летуче и может привлекать фуражиров к источникам пищи.
Еще по теме Передача информации о корме:
- 3.4.3. Использование генетической информации в процессах жизнедеятельности 3.4.3.1. Роль РНК в реализации наследственной информации
- 2.3.3. Поток информации
- Реализация генетической информации
- 5.5. ПУТИ ПРИОБРЕТЕНИЯ ОРГАНИЗМАМИ БИОЛОГИЧЕСКОЙ ИНФОРМАЦИИ
- ПРИЛОЖЕНИЕ Г Источники информации по пермакультуре
- 3.4.3.2. Особенности организации и экспрессии генетической информации у про- и эукариот
- Последующие этапы реализации наследственной информации
- ТОРФЯНЫЕ БОЛОТА - АККУМУЛЯТОРЫ И ИСТОЧНИКИ ПАЛЕОЭКОЛОГИЧЕСКОЙ ИНФОРМАЦИИ Н. К. Панова, Т. Г. Антипина
- 6.2. РЕАЛИЗАЦИЯ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ В ИНДИВИДУАЛЬНОМ РАЗВИТИИ. МУЛЬТИГЕННЫЕ СЕМЕЙСТВА
- ГЛАВА 6 ОНТОГЕНЕЗ КАК ПРОЦЕСС РЕАЛИЗАЦИИ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ
- 7.3. Самоузнавание и использование другой информации, полученной с помощью зеркала, у животных других видов
- Снова ламаркизм?
- 6-9. Горизонтальный перенос и генное единство природы
- ОСОБЕННОСТИ РАЗВИТИЯ ТРУТНЯ