Структура растительных сообществ
Поверхностная систематизация растительного покрова уже дает возможность осознать многообразие возможных подходов к анализу лежащих в его основе моделей и связей. В смешанном буковом лесу бросается в глаза структура популяции доминирующего бука, в которой чередуются всходы, молодые растения, доминирующие более старые деревья и, наконец, отмирающие (падающие от ветровала или поражения паразитными грибами) или вырубающиеся, а также упавшие гниющие стволы.
Подчиненный и мозаично распределенный в зависимости от небольших колебаний рельефа и почв или от световых условий нижний ярус состоит из разнотравья, граминоидов, мхов и лишайников (рис. 14.27). Геофиты, цветущие в весеннее время, такие, как Anemone nemorosa, летом и осенью полностью исчезают с поверхности земли, зато появляются плодовые тела шляпочных грибов, которые связаны с буком в качестве микоризных симбионтов или вместе с бактериями и мелкими беспозвоночными участвуют в процессах разложения листового опада. Другие подчиненные элементы биоценоза находятся на основаниях древесных стволов (листостебельные мхи) и на их коре (аэро- фильные водоросли и накипные лишайники).
Рис. 14.27. Части биоценозов (синузии) в центрально-европейском смешанном буковом лесу в конце лета (по F. Ehrendorfer):
А — впадины, заполненные листовым опадом, с относительно влажными нейтральными перегнойными почвами (с бактериями, грибами, мелкими беспозвоночными), по краям их — индикаторы влажности и соответственно богатства почвы: а — живучка ползучая (Ajuga reptans), b — лесная фиалка {Viola reichenbachiana)-, В — микроповышения, свободные от листового опада, с более сухими слабокислыми, бурыми лесными почвами, с наземной моховой и лишайниковой синузиями: с — гипновый мох (Нурпит cupressiforme), d — белый мох (Leucobryum glaucum), е — лишайник кладония {Cladonia pyxidata), f— микроповышения с плодовыми телами млечника (Lactarius vellereus; образует микоризу с Fagus) и с указывающими на кислотность и бедность питательными веществами почвы травянистыми розеточными растениями (гемикриптофитами), g — ожика беловатая {Luzula luzuloides), h — ястребинка лесная {Hieracium murorum), і — щучка извилистая (Deschampsia llexuosa): С — основание ствола с различными листостебельными мхами {Нурпит, Plagiothecium); D — кора бука с сину- зией эпифитных водорослей {Pleurococcus) и накипными лишайниками {Graphis scripta и др.); Е — древесный пень, заселенный грибами (плодовые тела трутовика Coriolus versicolor)
Подобная комплексная инвентаризация может быть проведена только путем разносторонних (и часто очень трудоемких) исследований.
На правильно выбранной пробной площади необходимо сначала охватить видовой состав и состав жизненных форм растительного сообщества, а также проанализировать его ярусное строение, чередование крупных и мелких биотопов и периодичность развития. Такие общие описания растительности и ее состояния впоследствии являются предпосылкой для выделения и описания растительных сообществ.Слово «структура» означает в немецком языке любую форму пространственного и иерархического подчинения или систему связей. Если в англоязычной специальной литературе речь идет о «community-» или «canopy structure» (структуре сообщества или покрова), то подразумевается в большинстве случаев структура геометрическая, в смысле «архитектуры». Применяемое в немецком языке обозначение видового состава растительного сообщества — струн тура насаждения = таксономическая структура соответствует аналогичному обозначению в английском «community composition» — состав сообщества.
Исходным пунктом для анализа растительности служит инвентаризация ее небольших выборочных образцов. Выбор и размер таких пробных площадей проводят в зависимости от того, какой биоценоз (или какая его часть) должен быть охарактеризован. Чтобы учесть все характерные деревья среднеевропейского леса, необходим минимум-ареал размером около 500 м2, а для богатых видами влажных тропических лесов площадь, возможно, должна быть около 1 га. Напротив, при описании лугов и дерновинных травяных сообществ достаточно уже 10—100 м2, а для сообществ мхов и лишайников — 0,1—4 м2. На альпийских лужайках на гранатных сланцах в северной части Скандинавии на площади около 1 м2 можно найти до 50 видов покрытосеменных. Если же площадь увеличить до 100 м2, то количество видов может возрасти до 60, а до 1 км2 — до 80. Таким образом, на площадке размером I м2 уже представлено 2/3 видового списка района. Наличие данных по зависимости числа видов от площади повышает ценность данных по биоразнообразию (например, по видовым числам). Диаграмма зависимости числа видов от площади (рис.
14.28) позволяет определить, с площади какого размера (минимум-ареала) можно получить достоверную выборку, содержащую более 95 % общего списка. При этом площади разной величины можно получать путем постепенного расширения стартовой площадки или же путем добавления других площадок, распределенных по обшей площади случайным образом. Показания эквивалентны не полностью. Как правило, второй метод объективнее и предпочтительнее.Для классификации растительных сообществ в целях определения типов сообществ (ранее это называлось фитосоциологией) применяется ряд параметров, среди которых выделяются две группы. Одна группа описывает характер произрастания особей видов в пределах пробной площади, которая должна по меньшей мере соответствовать величине минимум-ареала. Другая группа охватывает показатели, которые выводятся из сравнения большого числа таких площадей.
В пределах пробной площади можно определить или измерить: плотность особей, или обилие (число особей на единицу площади), участие в покрытии (% покрытия почвы вертикальной проекцией особей вида, так называемое доминирование в пространстве) и встречаемость (повторяемость встречи вида на площадках, заложенных внутри пробной площади, %). Высокая встречаемость более важных (обильных) видов указывает на высокую гомогенность растительного сообщества (в противоположном случае — гетерогенность).


?
Рис. 14.28. Минимум-ареал, или наименьшая площадь, которая содержит полный видовой набор растительного сообщества (gt; 95% всех видов)
Это можно выявить путем последовательного удвоения пробных площадей (А — метод одной площадки) или накопления добавочных данных, но на площадках такого же размера (В — метод многих площадок).
Результат выражен на кривой накопления (С), которая показывает, при какой величине пробной площади или при каком количестве повторяющихся площадок можно не ожидать существенного (gt; 5%) увеличения видового богатства. Типичный минимум-ареал при одной пробной площади для малопродуктивного луга или альпийской лужайки — 10—25 м2, для травяного яруса в лесах — 100—200 м2, для почти ненарушенного умеренного леса — 500— 1 ООО м2, для влажного тропического леса — более 1 га. Характер кривой зависимости видового числа от обследованной площади дает представление о гомогенности растительного сообществаРис. 14.29. Примеры распределения видов в растительном сообществе, которые находят отражение в различной встречаемости в пределах пробной площади:
А — регулярное распределение (случайно встречается в засушливых областях для «выдерживания дистанции», а также в моноклональных и антропогенных сообществах); В — случайное распределение (например, на неструктурированных рудеральных участках, в других случаях скорее редко); С — распределение группами или кучками (часто встречается на естественных лугах и в первичных лесах); D — очаговое распределение, ограниченное отдельными пятнами (англ. patches) часто при
точечных нарушениях или для клонов
К качественным признакам можно отнести общественность (социальность, произрастание группами или поодиночке) и распределение (регулярное или нерегулярное размещение; признак, близкий к встречаемости. рис. 14.29). В качестве меры оценки «успешности» вида (англ. vigor), с учетом его продуктивности, можно также применять индекс жизненности.
Из сравнения большого числа таких описаний можно определить константность вида (вероятность его встреч, как повторяющихся появлений на разных пробных площадях, аналог показателя встречаемости внутри пробной площади). Степень привязанности отдельных видов к определенным сообществам обозначают как их верность.
Кроме того, виды с очень высокой верностью (и обычно с более высоким обилием) типичны и называются характерными или характеризующими видами; другие, вполне типичные, также верные, но все же не столь строго связанные с этой ассоциацией, обозначаются как сопровождающие, а остальные — как случайные. Виды, которые делят сообщества одного ранга на подгруппы (например, субассоциации), называют дифференциальными, или разделяющими видами. Не будучи доминантами, они имеют сильную и специфическую сопряженность (верность) с соответствующей подгруппой и высокое постоянство в ее пределах, но не на ближайшей ступени более высокого социологического ранга. Дифференциальные виды, как правило, хорошие индикаторы определенных экологических условий.Чаще всего списки видов (так называемые перечни — releves) сопровождаются оценкой доминантности (покрытия) или плотности особей отдельных видов (доминантность и обилие объединяются в единый показатель мощности вида). Такая альтернатива возникает из-за необходимости разделять виды с узкими, вертикально направленными листьями и с широкими, плоско расположенными Виды, образующие листовые розетки, могут, несмотря на высокое покрытие, иметь сравнительно небольшую индивидуальную плотность. Некоторые злаки при высокой плотности побегов достигают сравнительно невысокого покрытия. У кло-
Таблица 14.2. Оценочная классификация значений обилия
|
Класс |
Покрытие, % |
Обилие |
|
5 |
gt; 75 |
Число особей любое |
|
4 |
50-75 |
Число особей любое |
|
3 |
25-50 |
Число особей любое |
|
2 |
5-25 |
Особи мелких форм роста, очень многочисленные |
|
1 |
lt; 5 |
Достаточно много |
|
+ |
Скудно |
Особи мелких форм роста, очень мало |
|
г |
Редко |
Почти единичные, редкие и вне пробной площадки |
нальных растений или растений, образующих большое число побегов, по чисто техническим причинам оцениваются не генетические индивиды (генеты), а побеги (раметы).
Значения обилия (= плотности) обозначаются обычно с помощью приблизительной классификационной шкалы (табл. 14.2).Эта ранговая шкала, составленная на оценочной основе, в практике часто для специальных целей еще дополняется или модифицируется в нижней области. По существу, она соответствует видоизмененным показателям оценки покрытия, благодаря которым доминирующие виды часто оцениваются несколько ниже, а редкие — выше. Точные количественные данные можно получить, только срезав всю растительность, пересчитав и взвесив все особи. Очень хорошая количественная оценка покрытия достигается методом «точечных квадратов», при котором в плотную, размеченную координатами рамку опускаются иглы и отмечается первое касание иглой особи одного вида в точке координатной сетки.
Таблица 14.3. Спектры жизненных форм (% участия соответствующих видов) некоторых важных формаций и экологических рядов (см. рис. 14.19, 14.42) (по С. Raunkiaer)
|
Область распространения |
Фанерофиты |
Хамефиты |
Гемикриптофиты |
Геофиты |
Терофиты |
|
Во всем мире |
46 |
9 |
26 |
6 |
13 |
|
От теплого до холодного (гумидного) климата |
|||||
|
Тропические дождевые леса |
96 |
2 |
2 |
||
|
Субтропические лавролист- ные леса |
66 |
17 |
2 |
5 |
10 |
|
Тепло-умеренные лиственные леса |
54 |
9 |
24 |
9 |
4 |
|
Холодно-умеренные хвойные леса |
10 |
17 |
54 |
12 |
7 |
|
Тундра |
1 |
22 |
60 |
15 |
2 |
|
От влажного до сухого (умеренного) климата |
|||||
|
Лиственные леса |
34 |
8 |
33 |
23 |
2 |
|
Лесостепь |
30 |
23 |
36 |
5 |
6 |
|
Степь |
1 |
12 |
63 |
10 |
14 |
|
Полупустыня |
59 |
14 |
27 |
||
|
Пустыня |
4 |
17 |
6 |
73 |
Каждое описание растительности снабжается данными, характеризующими местообитание, — это положение в рельефе, высота над уровнем моря, экспозиция, крутизна склона, тип почвы, характер материнской породы, хозяйственное использование и др. Постепенное или резкое изменение видового состава вдоль определенного градиента внешней среды (например, высоты над уровнем моря, влагообеспечен- ности. степени засоления, pH, светового довольствия) лучше всего может быть представлено в форме трансекты растительности (см. рис. 13.28). Она служит отправной точкой для по возможности наиболее объективного разграничения растительных сообществ. Часто хотя бы приблизительно ценность охарактеризованных признаков заключается в том, что, с одной стороны, эти признаки (особенно отношение покрытие/обилие) позволяют «взвесить» присутствие вида, с другой, благодаря большому количеству повторностей отдельные относительные неправильности все же не нарушают слагающейся достаточно четкой общей картины, это позволяет прибегать к количественным сравнениям.
Решающим признаком для общего габитуса каждого растительного сообщества являются морфология (форма) видов, а также их жизненные формы и формы роста (см. 4.2.4). По В. Раух (W. Rauh), форма роста— это принцип организации, или план строения, жизненная форма — это то, что может реализоваться от случая к случаю в жизненном пространстве в пределах того широтного пояса, условия которого это допускают. Форма роста дерева таким образом вполне может «спрессоваться» в определенных условиях среды в форму кустарника. Правда, разнииа в терминах в литературе почти не играет роли, оба понятия используются как синонимы (в немецкоязычной литературе это жизненная форма, в английской — форма роста, growth form). В разных зонах и основных типах растительности Земли существуют очень разнообразные жизненные формы (см. рис. 14.42, табл. 14.3), участие которых в сложении растительности обусловливает ее вертикальную стратификацию. Только самые первичные пионерные биоценозы, а также биоценозы, находящиеся в экстремальных условиях, имеют мало слоев (ярусов) или вообще однослойны. В большинстве естественных лесов есть:

Рис. 14.30. Циклическая регенерация горного елово-пихтово-букового девственного леса в Восточных Альпах (Ротвальд близ Лунца, 1 ООО м) (по K.Zukrigl, G. Eckhardt и J. Nather):
А — фаза омоложения с богатым подростом в «окнах» (места ветрового вывала); В — фаза оптимума с плотно сомкнутыми кронами и преобладанием хвойных пород; С — фаза разрушения перестой ного сообщества (много мертвых деревьев, как стоящих, так и упавших, повышается участие бука, вновь появляется подрост). Вертикальный и горизонтальный профили растительности: • ель (боковые ветви с темной заливкой); о пихта (боковые ветви без заливки); є бук (лиственные кроны изображены схематически); упавшие стволы, подрост— заштрихованы



• древесный ярус, часто с лианами и эпифитами, также разделяющийся на несколько ярусов;
- кустарниковый ярус, включая молодые деревья;
- травяной ярус с кустарничками и полукустарниками, включающий всходы деревьев;
- моховой, лишайниковый (напочвенный) ярус.
Этому наземному вертикальному расчленению растительности соответствует

Виды дубово-букового леса Древесный ярус
Carpinus betuius Quercus robur Fraxinus excelsior

Кустарниковый ярус
Crataegus spec.
Acer campestre Ilex aquifolium

Травяной ярус: весенне-зеленые растения
Anemone пет. + ranunc.
Ranunculus ficaria Corydalis cava Allium ursinum

Летне-зеленые растения, раннецветущие
Pulmonaria officinalis Mercurialis perennis Geum urbanum Phyteuma spicatum Melisa uniflora Poa nemorahs

Летне-зеленые растения, поздноцветущие
Campanula trachelium Circaea lutetiana Brachypcdium sylvaticum Galium sylvaticum

123456789 101112
Рис. 14.31. Сезонное развитие (фенология) характерных видов влажного дубово-букового леса в северо-западной части Германии (по H.EIIenberg).
Зимне-зеленые растения
Черным цветом показано развитие листьев этого года, горизонтальными линиями — перезимовавших листьев, вертикальными — цветков
Hepatica nobilis Oxalis acetosella Lamium galeobdolon Carex sylvatica Sanicula europaea Deschampsia cespitosa менее исследованная подземная ярусность корневых систем в ризосфере (см. рис. 13.24). Очевидно, что дифференцированное освоение воздушного пространства и почвенного слоя создает возможность для лучшего использования ресурсов (свет, влага, питательные вещества в почве, см. 14.2.4 2).
Традиционное деление растений на мелко и глубоко укореняющиеся очень упрощено и функционально плохо обосновано. Почти у всех многолетних растений имеются как поверхностные, так и глубоко проникающие корни, но их соотношение варьирует у разных видов и при этом сильно зависит от доступности влаги и питательных веществ. Небольшое число глубоких корней (которые часто не заметны) гарантируют растению даже во время критических периодов минимальное снабжение влагой (по меньшей мере для покрытия очень небольшого кутикулярного расхода влаги через устьичные отверстия), поверхностные корни обеспечивают усвоение большей части питательных веществ из особенно биологически активного верхнего слоя почвы. В периодически сухих районах глубина проникновения корней тесно коррелирует с сезонным ритмом активности побегов. Виды, которые остаются активными (зелеными) на время сухого сезона, имеют более глубокие корни, чем те, которые сбрасывают листву. На центрально-европейских умеренно сухих лугах, расположенных на известняках более 80 % всех корней располагаются в верхних 20 см почвенного профиля, хотя отдельные корни проникают вглубь до 6 м, что видно на отвесных обнажениях. Максимальную глубину корней более чем на 15 м можно наблюдать скорее как правило, нежели в виде исключения, в большинстве периодически засушливых областей Земли (табл. 14.3).
Растительный покров структурирован также и в горизонтальном направлении. В зависимости от величины свободной от растений поверхности различают открытую или сомкнутую растительность. Даже внутри насаждения, кажущегося однородным, часто можно наблюдать дифференцированное горизонтальное распределение видов в форме определенного рисунка, мозаики или коалиций (определенные виды группируются друг с другом или отстоят далеко, см. рис. 14.29). Уже незначительные повышения или впадины микрорельефа (см. рис. 12.13) обусловливают различия в снабжении влагой и питательными веществами (см. 13.6.2) и отсюда — в обилии видов. Сам по себе растительный покров также создает различные микробиотопы (см. рис. I4.27). Особое значение имеют окна в насаждении (англ gaps), образующиеся в результате выпадения особей, так как при этом возникает свободное пространство для поселения новых особей. Самое удачное типичное описание развития лесов ориентировано как раз на динамику в таких окнах (англ. gap dynamics, gap models). В нем речь идет прежде всего о продолжительности существования таких окон и сукцессии, которая в них развертывается. Рисунок 14.30 иллюстрирует цикл от возобновления (А) к оптимуму (В) и разрушению (С) лесного насаждения. Этот цикл зависит от длительности жизни доминирующих особей. В так называемых вырубочных окнах возобновление происходит не сразу, а протекает через стадии недолговечных пионеров, затем быстрорастущих, промежуточных (серийных) видов и только в последнюю очередь — молодых растений видов, изначально доминировавших (см. 14.3.2). В то же время закрепление молодых растений может происходить успешнее непосредственно под защитой других особей (англ. facilitation — облегчение; так, кактусы лучше поднимаются под защитой пустынных кустарников, что хорошо подтверждается на примере кактуса сагуаро — Carnegia gigantea).
Осень
Зима

ш Хамефиты ИВййЭ Геофиты
14
| Терофиты Гемикриптофиты
Р Нано-фанерофиты
^ Виды, не заметные над землей (покоящиеся)
Лето
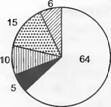
Наконец, сильно упорядочивающий/ структурирующий элемент— это еще и временная последовательность, периодизация, развития отдельных растений и сообществ в целом. Прежде всего важны такие фенофазы, как разворачивание листьев, цветение, плодоношение, старение листьев (рис. 14.31). В своей совокупности они обусловливают формирование в разные сезоны года различных «аспектов» (внешнего вида) растительного сообщества.
При сравнении сезонного развития видов влажных дубово-буковых лесов (см. рис. 14.31) отмечается, что листва на лиственных деревьях, более «открытых» для зимних морозов, разворачивается поздно. Вечнозеленым остается только Чех. Период до разворачивания листвы или во время разворачивания более благоприятен для цветения, особенно для ветроопыляемых (анемофильных) деревьев. Этот светлый
Рис. 14.32. Сезонные изменения участия растений разных жизненных форм в сложении растительности средиземноморской каменистой пустоши (Brachypodietum ramosi близ Монпелье, юг Франции). Проценты рассчитаны от общего числа видов (111) (по J. Braun-Blanquet)
период используют также требовательные к свету геофиты, которые развиваются только в короткое весеннее время. Остальные виды травяного яруса теневыносливы; произрастая непосредственно у поверхности почвы (защита от мороза), многие из них сохраняют листья в течение всей зимы. Для средиземноморской растительности в сезонных аспектах особенно характерно частичное или полное отмирание в летнее время терофитов (однолетников) и геофитов (луковичных и клубневых растений, рис. 14.32).
Итак, растительным сообществам свойственна пространственно-временная упорядоченная структура. Часто их большое видовое богатство и разнообразие жизненных форм и форм роста пространственно, функционально и сезонно связаны с наличием имеющихся экологических ниш, причем отношения между видами могут быть от взаимно дополняющих до зависимых и нарушения положительно влияют на сосуществование (см. 14.2.4.1).
Еще по теме Структура растительных сообществ:
- Изучение структуры сообществ
- 13.1.3 Свет в растительном сообществе
- Классификация морских донных растительных сообществ
- Пространственная структура растительности и биотопов
- Формирование и динамика растительных сообществ
- МИКРОСКОПИЧЕСКОЕ НАСЕЛЕНИЕ СФАГНОВЫХ БОЛОТ:СОСТАВ И СТРУКТУРА СООБЩЕСТВ Ю. А. Мазей
- РАСТИТЕЛЬНЫЕ СООБЩЕСТВА ДЛЯ ПЛОДОВОГО САДА В УСЛОВИЯХ УМЕРЕННОГО КЛИМАТА
- СТРУКТУРА РАСТИТЕЛЬНОГО ПОКРОВА НА ГРАНИЦЕЛЕС-ВОДОЕМ В ПОЙМАХ РЕК ОКСКО-ДОНСКОЙ РАВНИНЫ
- Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
- СОСТАВ СООБЩЕСТВ И КОМПЛЕКСЫ СООБЩЕСТВ
- Рифовые сообщества Особенности рифовых сообществ
- Гетеротрофные сообщества