Формирование и динамика растительных сообществ
Растительный покров находится в состоянии постоянного изменения (см. выше) и появляется на одних и тех же местообитаниях с различающимися в зависимости or сукцессионной фазы набором видов, спектром жизненных форм и доминантами.
Это хорошо видно из анализа, проведенного через длинные временные интервалы, и наблюдений на постоянных площадках длительного мониторинга (англ. permanent plots, permanent quadrants; рис. 14.33). Нехорошую картину происходящей сукцессии дают и сравнения различных участков сформировавшейся растительности на сходных местообитаниях (рис. 14.34).Реконструкция развития растительности в течение длительного времени проводится на основе ископаемых остатков (пыльцы) и свойств почвенного профиля (ископаемые почвенные горизонты и «горелые» горизонты). В семиаридных областях информацию относительно исторических смен доминирующих СЗ- и С4-растений можно проследить путем анализа соотношений изотопов углерода в гумусе (см. 13.7.4). Поучительны опыты, в которых можно наблюдать сукцессию после исключения нарушений (огораживание от травоядных животных, препятствование пожарам).
Для первоначального поселения и, соответственно, изменения видового состава требуются элементы расселения (см. диаспоры), привнесенные извне или имеющиеся в покоящемся состоянии в семенном банке почвы. Установлено, что на 1 м2 пахотной почвы содержится до 50 ООО жизнеспособных семян. В зависимости от зрелости системы в ней могут утвердиться популяции только определенных видов. На открытых участках с неразвитыми почвами (например, участки в зоне отступания ледников, галечные отмели, дюны) сначала формируется типичная пионерная растительность На нарушенных поверхностях (на мусорных участках, на обочинах дорог) вначале поселяются так называемые руде- ральные виды (см. рис. 14.13); для регулярно нарушаемых, используемых в сельском хозяйстве территорий характерна сегетальная флора (флора посевов и залежей).
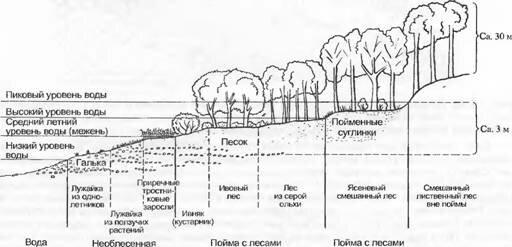
Каждая сукцессия связана с каким-либо изменением местообитания. Причины этого могут быть как внешними (аллогенная сукцессия), так и лежащими внутри самого сообщества (автогенная сукцессия).Так, постепенное становление пойменной растительности вдоль горных ручьев (рис. 14.35) после отложений на поверхности гальки, песка и ила (намыв берега) — аллогенная сукцессия. В стоячих водоемах (рис. 14.36), напротив, в образовании поверхности заселения преобладают органогенные отложения, образованные из самой растительности (зарастание), — это автоген-
1953
Рис. 14.33. Последовательная смена растительности (сукцессия) в течение 4 лет на площадке длительного наблюдения (1 м2), изначально не заросшей. Высохший торф верещатникового болота (Хил- ден, Рейнланд) (по S.Woike из R. Knapp):
а — Agrostis sp.; b — Molinia caerulea-, с — Sphagnum papillosum-, d — S. auriculatum-, e — Erica tetralix, f — Juncus bulbosus\ g — J. squarrosus; h — Dicranella cerviculata; і — Carex panacea; j, k — Eriophorum angustifolium: I — Cerastium sp.; m — Polygala serpyllifolia: n — Rhynchospora alba
Травяная пустошь "
| Кустарник —»-| Лес ¦
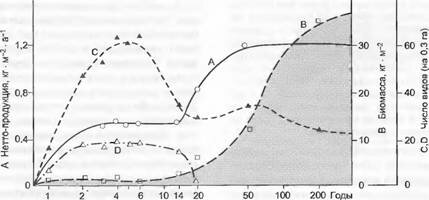
Рис. 14.34. Повторное зарастание лесом залежи в умеренной зоне (Северная Америка; Брукхэвен, Нью-Йорк) (по Holt и Woodwell из R. Н.Whittaker).
Через приблизительно 8 лет разнотравье и граминоиды сменяются летне-зелеными кустарниками, последние через 30 лет сменяются лиственными лесами, которые к 150 годам стабилизируются как кпимаксовый лес с летне-зелеными дубами и соснами. В ходе этой прогрессивной сукцессии увеличиваются первичная нетто-продукция (А, о—о) и биомасса растительного сообщества (В, о—о) вплоть до стадии климакса; напротив, падают число видов сосудистых растений (С, *—») после максимума в поздней фазе травяного сообщества и число адвентивных видов (D, д-д), элиминирующихся на кустарниковой стадии в результате конкуренции
из мягких (мелколиственных) древесных пород
из твердых (широколиственных) пород
Рис.
14.35. Схема последовательного распределения растительности в среднем течении реки в предгорьях Альп в зависимости от высоты воды и седиментации (наносов) (по Moor из H.EIIenberg)ная сукцессия. Из-за периодической стабилизации состояния среды (например, регулирование рек, относительно стабильное состояние водной поверхности вдоль скальных берегов) сукцессия может приостановиться, достигнув стабильного зонирования растительности. Аллогенные смены обусловлены изменениями климата или субстрата или нарушениями, автогенные управляются видами, имеющими особое значение в строении сообщества. В закреплении дюн это побегообразующие злаки, например Elymus и Ammophila (см. рис. 13.24; раздел 15.1.1), в зарастании водоемов — тростники и крупные осоки, в формировании среднеевропейских лесов — Fagus sylvauca. поскольку эти виды «затеняют» другие и в большей степени влияют на формирование органогенного горизонта почвы.
Атмосфера
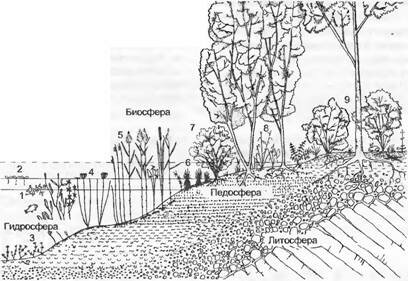
Рис. 14.36. Профиль растительности вдоль зоны зарастания эвтрофного озера в Центральной Европе (даны только названия родов, так как часто они представлены только одним широко распространенным видом в последовательности, соответствующей уменьшению глубины воды);
1 — свободно плавающие подводные растения, Utricularia (см. вставку 4.4, рис. В); 2 — свободно плавающие растения, Lemna (ряска), Hydrocharis (водокрас); 3 — прикрепленные подводные растения, Chara (вертикально растущая зеленая водоросль), Myriophyllum (уруть), Elodea (водяная чума, неофит), Hippuris (водяная сосенка); 4— прикрепленные водные растения с плавающими листьями, Nymphaea (кувшинка), Nuphar (кубышка), Тгара (чилим, водяной орех, в настоящее время очень редок), Potamogeton (рдест); 5— камыши (корневищные растения): глубже всех Schoenoplectus (= Scirpus, камыш), далее по степени обмеления Phragmites (тростник), особенно обычный в эвтрофных водах Typha (рогоз) и в мягкой воде на тростниковых торфах Sparganium (ежеголовка); 6 — пояс крупных осок с крупнокочкарными видами Carex sp., в болотных водах Menyanthes (вахта, трифоль), Potentilla palustris (сабельник) и с другими видами зоны зарастания, на краях болот Sphagnum (торфяной мох); 7, 8 — прибрежные деревья и болотистый лес, регулярно затопляемый, с Salix, Alnus и Populus; 9 — в лесу близ самой отметки уреза воды к элементам болотистого леса постепенно примешиваются элементы лиственного леса.
Отображены также основные сферы биотопа, причем пе- досфера и биосфера перекрываютсяЕдва растительность смыкается, в качестве движущей силы сукцессии вступают усиливающиеся взаимодействия между видами. Определенные виды больше не могут обновлять свои популяции, поскольку начинают доминировать другие. Они остаются в сообществе только в качестве стареющих реликтовых популяций, в то время как следующая фаза сукцессии уже дает о себе знать появлением молодых растений подроста. Типичные примеры — береза и сосна в смешанных лесах. Они представляют собой в таких лесах чаще всего стареющие реликты более ранних стадий сукцессии (и таким образом являются показателями бывших нарушений) и как светолюбивые деревья не могут больше возобновляться при полной сомкнутости крон. Только в заключительной стадии развития растительности (климаксовая растительность) возникает относительно стабильное равновесие в видовом составе (см. рис. 14.34), а также между размножением и отмиранием участвующих в сообществе видов. Полностью стабильное состояние все же не достигается никогда. «Зрелые» растительные сообщества, которые никогда не были вовлечены в хозяйственный оборот и не испытывали катастрофических нарушений (сплошная рубка, использование под пастбища, пожары, затопление), всегда представляют собой мозаику разных сукцессионных стадий.

Естественное развитие
Лесное хозяйство в низкоствольных лесах Сплошная рубка
Нерегулярный покос или выпас
— — Регулярный выпас с удобрением
Регулярный сенокос с удобрением
Вспашка
Рис. 14.38. Динамика растительности Центральной Европы под воздействием хозяйственного использования: регрессивные и (вторично) прогрессивные сукцессии на глубоких пологих известняковых склонах у подножия Высокого Венна (ФРГ). Растительные сообщества связаны в «кольца» соответствующих комплексов растительности и занимают определенную «плиту» (по M.Schwickerath)
В бореальной зоне и верхнем поясе гор влажно-умеренного климата Северного полушария каждая автогенная не нарушаемая сукцессия происходит в направлении хвойного леса, на плоских равнинах — лиственного леса.
Различают первичную сукцессию на свежих, только что освободившихся поверхностях (после отступления ледника, изменения русла реки) и вторичную сукцессию, например, на заброшенных залежах или после недавнего пожара. Соответственно чему вошло в практику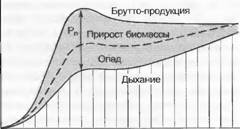
Фаза развития Фаза зрелости Фаза старения
Возраст
насаждения
Рис. 14.37. Фазы развития однородного лесного насаждения.
Пропорции дыхания, опада (VA), прироста биомассы (АВ), нетто- (Рп) и брутто-продукция; отчуждение травоядными животными не принимается во внимание (по Т. Kira и Т. Shidei)
различать леса первичные и вторичные, это
не означает, что первичный лес никогда не нарушался естественным образом, но такие нарушения остались далеко позади. За очень долгое время (для лесов более сотни лет) сукцессионные ряды сходятся, и влияние макроклимата становится все более преобладающим, в то время как другие факторы отходят на второй план (зональная растительность). В Центральной Европе за очень небольшими исключениями существуют только вторичные леса различной степени «естественности» (обратно пропорционально степени окультуривания = гемеробии). В ландшафтах, никогда не использовавшихся человеком, ту стадию развития, на которой сукцессия приостанавливается, предрешает частота естественных нарушений (горные осыпи, лавинные полосы, зоны затопления, дюны, пожары, инвазия травоядных и др.).
Зональные типы растительности могут выступать так же, как и экстразональные, за пределами своей исконной области распространения, если локальный климат соответствует макроклимату этой основной области (например, произрастание субмеридиональных лесов из дуба пушистого на сухих южных склонах в Западной Европе).
Экосистема может меняться тем быстрее, чем меньше ее биомасса (В) и чем интенсивнее в ней потоки вещества и энергии.
Если прирост биомассы на единицу времени обозначить как ДВ, то отсюда следует, что длительность полного оборота (turn-over) биомассы В/ДВ = 1. Сообщества планктона или терофитов могут соответственно этому изменяться за дни или месяцы, лесные же сообщества — за десятилетия и столетия.При формировании одного однородного растительного сообщества можно параллельно сменам стадий развития, зрелости и старения установить специфические изменения В, ДВ, дыхания (R) и продуктивности (Рп) насаждения (рис. 14.37). Эти изменения связаны с тем, что с прогрессирующим возрастом насаждения соотношения автотрофных компонентов (листьев) и гетеротрофных компонентов (стволов, ветвей и корней) все более уклоняются в сторону гетеротрофных и насаждение наконец угасает, если не происходит его омоложения.
В ходе прогрессивной сукцессии комплексных биоценозов (см. рис. 14.33) также сначала происходит прирост В и Р„, так как ДВ больше, чем VA + Ук. Такая экосистема продуктивна, но еще относительно изменчива и нестабильна. На стадии климакса наконец достигается фаза естественного омоложения, которая обеспечивает стабилизацию В на высоком уровне, поскольку прирост ДВ расходуется в ходе биологического круговорота. Отношение Рп и VA + VK, так же как и опада и разложения опавшего материала, уравнивается. Такая экосистема называется протективной. Как и климаксовая экосистема, она остается достаточно стабильной и имеет максимальную биомассу с очень небольшим нетто-приростом.
Регрессивные сукцессии идут в сторону от климаксовой растительности и связаны с ее деградацией. Если не принимать во внимание природных катастроф (землетрясения, ураганы и пр.) или катастрофических биологических изменений (например, голландская болезнь вязов, см. Ophiostoma), они почти всегда являются реакцией на вмешательство человека (см. рис. 13.51). Самыми тяжелыми являются те вмешательства, которые непосредственно или через ослабление растительности наносят ущерб почвам (перевыпас, регулярное выжигание).
Природные и антропогенные смены растительности на известковых почвах нижних горных поясов Центральной Европы обнаруживают как прогрессивные, так и регрессивные тренды, которые, будучи циклически связаны между собой (рис. 14.38), все же обратимы. Регрессионные смены, приводящие к ухудшению почв, такие, как перевыпас и повышенная частота пожаров на каменистых пустошах, ведущие к деградации средиземноморских дубовых лесов, в значительной степени необратимы.
Еще по теме Формирование и динамика растительных сообществ:
- Разложение растительных остаткови формирование подстилки
- РАЗЛОЖЕНИЕ РАСТИТЕЛЬНЫХ ОСТАТКОВ И ФОРМИРОВАНИЕ ПОДСТИЛКИ
- 13.1.3 Свет в растительном сообществе
- Классификация морских донных растительных сообществ
- Структура растительных сообществ
- СОСТОЯНИЕ И ДИНАМИКА ЛЕСНОЙ РАСТИТЕЛЬНОСТИ НА ЕСТЕСТВЕННЫХ БОЛОТАХ БЕРЕЗИНСКОГО ЗАПОВЕДНИКА
- РАСТИТЕЛЬНЫЕ СООБЩЕСТВА ДЛЯ ПЛОДОВОГО САДА В УСЛОВИЯХ УМЕРЕННОГО КЛИМАТА
- ДИНАМИКА ЛЕСНОЙ РАСТИТЕЛЬНОСТИ ИБОЛОТООБРАЗОВАТЕЛЬНОГО ПРОЦЕССА В ГОЛОЦЕНЕВ ГОРАХ СРЕДНЕГО УРАЛА
- Продуктивность сообществ и жизненные стратегии водорослей. Морфофункциональный подход к изучению сообществ макрофитов
- СОСТАВ СООБЩЕСТВ И КОМПЛЕКСЫ СООБЩЕСТВ