ЛОЛИЭМБРИОНИЯ
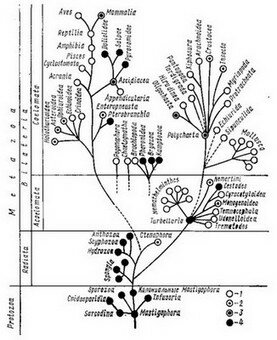
Рис. 146. Распространенность бесполого размножения в животном мире (т О. М. Ивановой, 1970):
1 — классы, в которых бесполое размножение отсутствует, 2 — встречается редко, 3 — свой ствеиио целым семействам и отрядам, 4 — свойственно всем представителям
подчеркнуть, что у Oceania не все бластулы делятся, это не обяза тельная полиэмбриония. У некоторых кишечнополостных могу «почковаться» планулы (Chryosaora, Alcyonaria); у Lumbriculu trapezoides может иметь место полиэмбриония на стадии бластуль Закономерно наступающие в ходе развития случаи полиэмбри
«нии характеризуют некоторых насекомых, особенно наездников (Encyrtus, Polygnotus, Xithomastis и др.).
Рассмотрим в каче- отве примера зародыш паразитического наездника Ageniaspis fus- .ciollis, откладывающего яйца в кладки насекомого Hyponomeuta. Происходит развитие зиррулоподобных групп клеток (по 12—15 клеток в каждой группе, а иногда и до 60—80). Морулы могут, в свою очередь, разделиться на две группы, каждая из которых может развиться в самостоятельный организм (рис. 147).
Хорошо известна полиэмбриония у мшанок. Морулообразный зародыш (рис. 148,Л) принимает неправильную лопастную форму (рис. 148,Б) и затем почкуется, давая начало многим вторичным зародышам (рис.
148,В). Вероятно, еще не все случаи полиэмбрионии описаны. Потенциальные возможности полиэмбрионии у многих животных могут быть доказаны, например, результатами опытов по изоляции бластомеров, по экспериментальной полиэмбрионии у амфибий и птиц, о чем говорилось в гл. XI.
Даже у млекопитающих встречается или закономерно наступающая полиэмбриония (например, у броненосцев), или вызываемая экспериментально. К случаям полиэмбрионии можно отнести и развитие однояйцевых однополых близнецов у человека, несомненно развивающихся из частей раннего эмбриона.
У броненосцев «делится» бластоциста, возникают не одна (как обычно), а несколько зародышевых полосок, из которых развиваются новые зародыши (у Tatusia hybrida — 8, у Т. novemcincia их 4). Новые эмбрионы имеют общую плаценту при самостоятельном для каждого зародыша амнионе (рис. 149).
МОРФОФИЗИОЛОГИЧЕСКАЯ ОСНОВА ПРОЦЕССОВ БЕСПОЛОГО РАЗМНОЖЕНИЯ И ОСОБЕННОСТИ РАЗВИТИЯ ПРИ БЕСПОЛОМ РАЗМНОЖЕНИИ
Разнообразие форм бесполого размножения у животных разной сложности и своеобразия организации очень велико; соответственно своеобразны и морфогенетические процессы, сопровождающие бесполое размножение, а потому нелегко установить общие биологические закономерности. У ряда животных бесполое размножение возникло как вторичное явление.
Наступление бесполого размножения связано либо с нарушением нормальной интеграции организма, либо тех или иных его систем.
О. М. Иванова-Казас (1976) доказывает, что имеется принципиальное сходство закономерностей индивидуального развития при бесполом и половом размножении. Анализируя явления бесполого размножения у Cnidaria, Bryozoa, Tunicata, автор считает, что к бластогенезу применимы такие понятия, как «рекапитуляция», «филэмбриогенез», «палингенез», «ценогенез» и др. С этим трудно согласиться. Решающим является факт, что исходная клетка при эмбриогенезе — зигота, а при любой форме бластогенеза и соматического эмбриогенеза — это соматические клетки, входившие до начала размножения в состав определенных органов и тканей материнского организма-, т.
Некоторые исследователи (Р. Бриен, 1956) утверждают, что бластогенез — это совершенно новый, более поздний в филогенетическом смысле тип развития, возникший без связи с эмбриогенезом и имеющий другие закономерности. Однако это противоречит тому, что большинство просто организованных животных размножается бесполым путем.
При бесполом размножении дезинтеграция может затрагивать весь организм или происходить локально. Паратомия v Oligochaeta состоит в том, что в результате каких-то внутренних процессов (без повреждения внешними факторами) и определенной части тела появляется зона деления. В этой зоне происходит частичный некроз клеток, изменяются клетки наружного эпителия, эпителия кишечника, клетки мезодермального происхождения, происходит де- Дифференциация эктодермальных и мускульных клеток. Прежние структуры тканей в зоне деления радикально изменяются. П. П. Иванов, в 1903 и 1907 гг. тщательно изучавший регенерацию и бесполое размножение у Lumbriculus и других олигохет, сделал заключение: при делении одной особи на две организм должен пройти момент временного нарушения нормальных физиологических связей между частями тела, а этот момент сопровождается отмиранием одних клеток, дедифференцировкой других. Таким образом происходит локальная илн относительно локальная дезинтеграция тканевых систем.
Вероятно, состояние местной дезинтеграции может наступать и: при тех или иных формах почкования, например при почковании гидры. Б. П. Токин (1959) предполагает, что в зоне почкования нарушаются нормальные морфофизиологические взаимоотношения между клетками в связи с процессами пищеварения. Особенно очевидная дезинтеграция или дезорганизация тела животного, сопровождающая бесполое размножение, на примере губок. При гемму- лообразовании большая часть тела губок распадается и без того- мало интегрированные организмы нацело теряют свою нормальную организацию.
О. М. Иванова-Казас (1970, 1976) говорит о морфофизиологических причинах наступления бесполого размножения и лишь в отношении некоторых случаев находит возможным применить термин «дезинтеграция». Действительно, далеко не всегда имеют место видимые процессы, предшествующие бесполому размножению,, которые можно было бы обозначить словом «дезинтеграция» тканей. Одиако при любом типе бесполого размножения требуется выяснить, как и почему наступает развитие новой особи из клеток,, принадлежащих до момента почкования или деления материнскому организму.
Почкование асцидии Clavetina связано с явлениями местной (летом) или общей (осенью) дезинтеграции; новый организм строится заново из соматических клеток; никакой преемственности: между полярностью материнского организма и таковой почки нет.
Многие асцидии обладают способностью к «регенерации», точнее — к соматическому эмбриогенезу. Экспериментаторы ие раз; убеждались, что практически любой участок тела асцидий можег дать целый организм и что развитие новых особей может происходить из участков тела, лишенных производных того или иного зародышевого листка или даже состоящих из клеток только одного зародышевого листка.
В заключение следует отметить, что, вероятно, всегда развитие новой особи в результате бесполого размножения происходит не из одной соматической клетки, а из групп клеток, которые можно представить себе как подобие зародышей на стадии бластулы. Впрочем, вопрос этот еще далек От ясности. Так, некоторые исследователи предполагают, например, что клетки почек медуз являются продуктом деления одной омоложенной клетки (Б. Брам, 1908, 1911).
Источник: Токин Б. П.. Общая эмбриология: Учеб, для биол. спец, ун-тов.—4-е изд., перераб. и доп. 1987
Еще по теме ЛОЛИЭМБРИОНИЯ:
- Портал "ПЛАНЕТА ЖИВОТНЫХ". Кто ты, собака?, 2010
- Любопытное доказательство того, что собаки очень давно одомашнены, приводит советский ученый-языковед академик Н. Я. Марр...
- Антропологи изучают кости и скелеты людей очень далекого прошлого, изучают их близких и отдаленных родственников — ископаемых и современных обезьян,— чтоб восстановить путь, который прошел человек в своем развитии.
- Находки, проливающие свет на происхождение собак, имеют возраст 8—10 тысяч лет...
- Значит, собака пришла сюда вместе с человеком?..
- Находка Савенкова произвела сенсацию...
- КАК ЭТО МОГЛО СЛУЧИТЬСЯ!
- Люди не очень опасались волков, волки же не очень боялись людей и нередко подходили к стоянкам первобытного человека достаточно близко.
- Видимо, много, очень много лет жили люди и волки на расстоянии, очень медленно сближались и очень трудно понимали выгодность сближения...
- Среди ученых нет единого мнения, ради чего была приручена собака...
- Но могло быть и иначе.
- Волки жили стаями...
- Cуществует еще одна гипотеза, объясняющая появление волков среди людей...
- ПОЧЕМУ ЭТО МОГЛО СЛУЧИТЬСЯ!
- Как относились к животным наши далекие предки?..
- "УМОМ СОБАКИ ДЕРЖИТСЯ МИР"