Солёность
Жизнь существует в водоёмах с солёностью от 0 до примерно 300%о. Однако распределение вод с разной солёностью по этому диапазону существенно неравномерно. Большая часть водоемов суши имеет солёность близкую к пресной.
Моря и океаны имеют солёность около 35%о, её называют нормальной океанической. В настоящее время солёность в Мировом океане колеблется в очень небольших пределах: в аридных районах может возрастать до 39%о (восточная часть Средиземного моря) и даже 41%о (Красное море), в местах таяния льда в высоких широтах — может опускаться до 32-33%о. Во внутренних морях и местах впадения рек солёность может быть существенно иной, но относительная площадь таких акваторий очень незначительна. Солёность океана Земли была примерно постоянной как минимум на протяжении фанерозоя* и, поскольку моря Земли всегда были фактически частями одного Океана, по-видимому никогда не было существенных географических различий ни в общих значениях солёности, ни в соотношениях разных элементов. Поэтому на Земле сформировались две основные водные биоты: пресноводная и морская. В редких случаях длительно существующих крупных бассейнов с отличающейся от морской солёностью (например, Каспий и связанные с ним водоёмы), в общем формировались те же биоты с некоторой местной спецификой (подробнее см. ниже). Различия морской и пресноводной биот между собой почти такое же, как каждой из них с наземной. Наземная и пресноводная биоты несомненно произошли от морской, при этом не было создано таксона животных рангом выше класса. Морская биота гораздо разнообраз-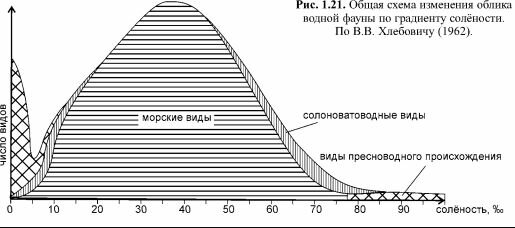
* Существенное изменение химического состава морской воды произошло только при появлении свободного кислорода. В этот период произошло окисление железа, марганца и других металлов и выпадение их солей. Именно с этим периодом связано возникновение крупных железорудных месторождений (например, Курского).
нее пресноводной. Общее число типов в море — 34, в пресных водах — 17. Число классов в море — 73, в пресных водах — 35. Для морской биоты эндемичны радиолярии, некоторые кишечнополостные (сифонофоры, сцифомедузы, коралловые полипы), гребневики, приапулиды, сипункулиды, эхиуриды, погонофоры, большая часть классов моллюсков (Monoplacophora, Aplacophora, Polyplacophora, Cephalopoda, Scaphopoda), пантоподы, щетинкочелюстные, иглокожие и оболочники. Почти исключительно морскими являются гидромедузы, немертины, полихеты и элазмобранхии. Но и пресноводная биота имеет свою специфику: солнечники, коловратки, олигохеты, пиявки, кладоцеры, насекомые, мшанки Phylactolaemata, жаброногие раки, двоякодышащие рыбы и амфибии обитают исключительно или почти исключительно в пресных водах. Видов, которые обитали бы как в морской, так и в пресной воде немного. Более того, детальное рассмотрение их биологии показывает, что наиболее ответственные стадии их жизненного цикла (размножение и раннее развитие) проходят или только в пресной, или только в морской воде. Видов, способных размножаться и там, и там практически нет. Столь чёткое разделение морской и пресноводной биот является фундаментальным свойством жизни на Земле и проявляется на разных уровнях организации: экосистемном, организменном, клеточном и молекулярном. Т. е. солёность водоёмов влияет на функционирование живых систем на уровне экосистем, физиологии и биохимии (Хлебович, 1974).
Локально солёность может иметь другие значения. Большая часть таких отклонений наблюдается в прибрежных районах. Чаще всего встречается распреснение под влиянием материкового стока. При смешивании пресной и морской воды солёность меняется плавно. Однако ответ биоты, как это обычно бывает, дискретен. Это означает, что до определённого предела биота как бы не замечает изменения солёности. При дальнейшем изменении солёности происходит резкое изменение биоты. В зоне контакта пресных и морских вод наблюдается падение видового богатства и биомассы.
Самая существенная перестройка биоты происходит на стыке морской и пресноводных биот и приурочена к солёности 5-8%о (рис. 1.21). Менее выраженные границы локально выявляются и при других солёностях. В соответствии с этими рубежными значениями было предложено несколько классификаций. Приуроченность биот (и таксонов, из которых они состоят) к разной солёности описывают терминами, образованными обычными приставками (олиго-, мезо-, гипер-, стено- и поли-) и корнем -галинный. Олигогалинными называют организмы, встречающиеся при низкой солёности (но не пресноводные), мезогалинными — встречающиеся при средней, полигалинными — при океанической, гипергалинными — при солёности существенно выше океанической. По широте диапазона, при котором встречаются таксоны, выделяют стеногалинные — встречающиеся в узком диапазоне солёности, и полигалинные — встречающиеся в широком диапазоне солёности.В общем, при возрастании отклонения солёности от океанической начинается прогрессирующее выпадение различных таксономических групп. Наиболее бросающимся в глаза является уменьшение числа видов одной из ведущих в донных сообществах групп — иглокожих. Так, в Баренцевом море (океаническая солёность) отмечено почти 100 видов иглокожих, а в Белом (солёность 25-30%о) — всего 22, причём многие виды, обильные по всему Баренцевому морю, в Белом очень редки. В Чёрном море (солёность 18%о), несмотря на его гораздо более южное расположение (видовое разнообразие большинства таксонов макробентоса растёт к экватору), из иглокожих в массе повсеместно встречается только один эндемичный вид — офиура Amphiura stepanovi, местами обильны голотурии и только сейчас происходит распространение из Прибосфорского района морской звезды Asterias rubens.
При дальнейшем уменьшении солёности (ниже 5-8%о) локальное видовое разнообразие вновь растёт, поскольку начинают появляться виды пресноводной биоты.
При впадении рек в океан переход пресных водных масс в морские обычно совершается быстро, формирующиеся при этом мезогалинные и олигогалинные водные массы непостоянны во времени и в пространстве и занимают малые площади, в связи с чем специфическая мезогалинная и олигогалинная биоты не развиваются.
Олигогалинные и мезогалинные водные массы населяют эвригалинные виды, как морского, так и пресновод- ного происхождения, и виды, называемые солоноватоводными. В действительности, большая часть солоноватоводных видов — это также эвригалинные виды, как морского, так и пресноводного происхождения, но, из-за малой конкурентоспособности в морских и пресных водах, они встречаются преимущественно в солоноватых водах.га ш ш
1 2 3
0 2 4 6 8 10%о
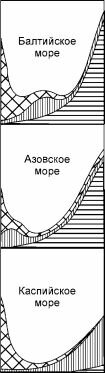
Рис. 1.22. Схема изменения числа видов по градиенту солёности в различных морях. Обозначения как на рис. 1.15. По JI.A. Зенкевичу (1959).
Например, по нашим данным (Днестровская, Жирков, 2008), полихеты Micronephthys minuta и М. neotena обитают в водоёмах как с океанической солёностью, так и в солоноватоводных. Однако в акваториях с океанической солёностью они встречаются гораздо реже. Так, в российских коллекциях из открытых районов Баренцева моря имеется всего 4 экземпляра М. minuta, а М. neotena отсутствует вовсе. В то же время в солоноватом Белом море плотность их поселений достигает 6000 экз./м2. Ещё более показателен пример полихет Spionidae (Сикорский, 1992). Спиониды питаются, собирая детрит с поверхности грунта. Организмы с таким типом питания обитают преимущественно на рыхлых грунтах (ил, глина и т. п.). В полном соответствии с этим в Белом море арктические шельфовые виды Spio theeli и два вида рода Marenzellaria довольно обычны на рыхлых заиленных грунтах, богатых детритом. Однако в Баренцевом море в районах с нормальной океанической солёностью они встречается исключительно на чистых крупнозернистых хорошо промытых песках, где практически отсутствуют другие виды с аналогичным типом питания. Два оставшихся арктических шельфовых вида спионид (Scolelepis burkov- skii, Spio petersenae) известны по единичным находкам, но также с крупнозернистого промытого песка из районов с нормальной солёностью.
Такое распространение невозможно объяснить малой способностью видов к расселению. Единственное возможное объяснение — малая конкурентоспособность арктических шельфовых видов. Поэтому они встречаются в маргинальных биотопах, куда их вытесняют другие виды со сходным типом питания, входящие в состав арктической шельфовой биоты.В целом для олиго- и мезогалинных вод характерно общее обеднение видового состава. Специфическая биота не образуется даже в таких сравнительно крупных существенно распреснённых водоемах, как Балтийское, Чёрное и Азовское моря, хотя отдельные эндемичные виды и возникают.
Принципиально иначе описывают ситуацию в Понто-Каспийском бассейне (Зенкевич, 1959; Мордухай-Болтовской, 1960; Хлебович, 1974 и др.). В своей истории Понто- Каспийский бассейн и отдельные его части неоднократно испытывали периоды осолонення и опреснения. При этом связь с океаном длительное время отсутствовала, в связи с чем в нём сформировалась эндемичная Каспийская биота. Эту биоту рассматривают как единую солоноватоводную, противопоставляя её другим биотам, обитающими совместно с ней: средиземноморской, арктической и пресноводной. В действительности тут происходит смешение и подмена понятий: разные по происхождению группы видов отождествляют с разными биогеографическими (т. е. распространёнными по-разному) группами. Это действительно сильно перекрывающиеся, но вовсе не идентичные понятия. Эта подмена приводит к искажённому описанию экосистем данного региона. Наиболее существенное искажение происходит при рассмотрении эндемичной биоты как единого целого (рис. 1.22). Безусловно, по происхождению эта биота едина, однако отнесение её к единой солоноватоводной основано лишь на трёх фактах:
- Понто-каспийские виды не переносят солёности близкие к океанической, и могут нормально существовать лишь при солёности, которую в условиях полносолёных морских водоемов следует считать солоноватой.
В действительности этот факт свидетельствует лишь о том, что понто-каспийские виды адаптировались к условиям, существующим в их ареале.
Да, если бы они обитали при такой солёности за пределами этого ареала, их можно было бы назвать солоноватоводными, но за его пределами обитают отнюдь не все экологические (различающиеся со- лёностными предпочтениями) группы.- За пределами Каспийского (а ранее — и Аральского) моря (в первую очередь в Чёрном и Азовском морях) каспийские оп происхождению виды обитают в солоноватых водах, преимущественно при солёности ниже 8%о: в эстуариях, в прошлом бывших частью Понто-Каспийского бассейна, или имевших связь с ним, а ныне являющимися рефу- гиумами.
В действительности виды, обитающие в Азово-Чёрноморском бассейне, живут и в Каспии (или там живут крайне близкие формы). Однако, если посмотреть на их распространение в Каспии, то окажется, что и там эти виды живут в солоноватых водах. Лишь немногие из них распространены по всей акватории Каспия, но в целом они предпочитают солёность ниже 8%о (например, Hypania kowalewskii — Голикова и др., 1987). Кроме этих видов, в Каспии обитает другой комплекс видов, обитающий преимущественно или исключительно в Среднем и Южном Каспии при солёности выше 8%о. Ни один из них в Азо- во-Чёрноморском бассейне не встречается.
- С точки зрения гидрологии и гидрохимии (т. е. по содержанию солей) каспийская вода действительно солоноватая.
Однако характеризуя биоту, надо исходить из признаков биоты: характеристику видов (морской или солоноватоводный) надо давать не по абсолютным значениям солёности, а по распределению их по градиенту «река - нормальная солёность водоёма», в которую она впадает. В результате классифицирования видов по признакам их абиотического окружения, а не по их реальному распространению, к солоноватоводной биоте относят не только виды, действительно обитающие в солоноватых водах эстуариев и лиманов Пон- то-Азова и при той же солёности в Каспии, но и виды, которые в реальности в солоноватых водах (т. е. при солёности ниже 8%о) не обитают или, по крайней мере, их не предпочитают, а лишь способны там существовать (т. е. достаточно эвригалинные).
Изучение распределения отдельных видов вдоль градиента солёности показывает, что понто-каспийские виды, в том числе и автохтонные, биогеографически относятся не к одной солоноватоводной биоте с уникальными свойствами и неясными солёностными предпочтениями, а к трём обычным, распространение которых совершенно различно, из- за чего относить их к одной биогеографической группе никак нельзя:
- Пресноводная биота — характерна для пресной воды. В её состав биоты входит большое число каспийских по происхождению видов, которые широко распространились по рекам, в первую очередь бассейну Волги. Из бентоса — это двустворчатые моллюски (.Dreissena polymorpha и, отчасти, D. bugensis), бокоплавы (26 видов), полихеты (Hypania invalida и Н. kowalewskyi) и др. Из планктона — это хищные кладоцеры. К этой же группе относится не менее 18 видов рыб (в первую очередь бычки, а также осетровые, некоторые виды которых выходят в море для питания, но все они размножаются только в пресной воде). Многие виды этой группы проникают в Северный Каспий.
- Солоноватоводная биота— характерна для вод с низкой солёностью в Каспии и Азово-Черноморском бассейне: Monodacna caspia, Dreissena polymorpha andrusovi, Hypanis vitrea, H. colorata. Виды этой группы или общие для Каспия и Азово-Чёрномор- ских лиманов, или в этих двух районах обитают эндемики. Н. colorata недавно вселилась в Каспийское море из эстуариев Азовского, в Каспийском море она населяет только его Север, где солёность та же, при которой этот вид встречается и в Чёрном и Азовском морях.
- Морская биота— характерна для солёности, наблюдающейся в Среднем и Южном Каспии (обычно более 8%о), в Азово-Чёрноморском бассейне виды этой биоты не ветре- чаются. Сюда относятся из каспийских эндемов: двустворчатые моллюски: Didacna trigonoides, D. profmdicola, D. longipes, D. baeri, D. protracta, Hy- panis albida, H. caspia, H. semipellucida, Dreissena rostriformis и др., 69 видов брюхоногих, полихета Hypania brevispina, 5 видов мизид, 18 видов гаммарид, 1 корофиида (Corophium spinulo- sum), 4 вида кумовых, 16 видов нематод, 1 вид губки, мшанка (Victorella pavida). В состав этой биоты вошли и недавние вселенцы из Средиземного моря (Cerastoderma glaucum, С. rhomboides, Zostera папа и др., а также вселившиеся в XX в. Mytilaster, Abra, Nereis и др.) и виды так называемого арктического комплекса.
«в Понтоазове... почти совершенно отсутствуют виды, обитающие только в Среднем и Южном Каспии» (Мордухай-Болтовской, 1960: 63).
Т. е. распределение биот по градиенту солёности в Каспии и шире — в Понто-Кас- пии такое же, как в остальном Мировом океане, с тем отличием, что здесь имеется гораздо более богатая, чем обычно, солоноватоводная биота, а морские виды приспособлены к обитанию в воде с солёностью существенно ниже океанической (но той, которая свойственна их ареалу!). В Каспии при солёности 3-8%о наблюдается тот же минимум видового богатства и биомассы морских видов. Понто-Каспийских видов там немного, их наибольшее разнообразие в Каспии наблюдается при солёности gt;10%о, где и в Мировом океане появляются в значительном количестве морские виды, а солоноватоводные исчезают.
Казалось бы, казуистика. Однако уменьшение черт уникальности Каспия делает установленные закономерности по смене биот вдоль градиента солёности более универсальными и позволяет понять их глубже. Выделение трёх биот в Каспии важно и для лучшего понимания биологической структуры Каспия в целом и для природоохранных целей в частности. Кроме того, такая интерпретация позволяет понять, почему в Каспии зона минимума заполнилась Nereis и Abra — морскими, казалось бы, видами. Почему при вселении в Каспий видов из полносолёных морей происходит не вытеснение каспийских видов в солоноватые районы (как произошло бы, если каспийские морские виды были бы действительно солоноватоводными), а их совместное обитание или даже вымирание каких-то видов (как понто-каспийских, так и вселенцев).
Колебания солёности в Понто-Каспии привели к формированию самого богатого в мире комплекса солоноватоводных и богатого — пресноводных организмов, булыпей частью морского происхождения. Поскольку морская биота гораздо разнообразней пресноводной, многие из этих видов оказались весьма конкурентоспособными, и при возникновении в последнее время возможности для расселения за пределы рек Понто-Каспийско- го бассейна быстро расширяют свой ареал.
Также колебания солёности в Понто-Каспии привели к снижению общего разнообразия морской каспийской биоты, появлению в ареале экосистем неиспользуемых ресурсов, которые стали потреблять организмы пресноводного происхождения. Так, основные хищные рыбы Каспия — осетровые (размножаются в пресной воде). В зоопланктоне Северного (но не Южного и Среднего) Каспия существенную роль играют Cladocera, роль которых в зоопланктоне Мирового океана очень незначительна (подробнее об особенностях каспийской биоты см. ниже, гл. «Биогеография моря»).
С увеличением солёности выше океанической происходит снижение видового богатства. Здесь можно выделить гипергалинные воды (диапазон примерно 45-80%о) и уль- трагалинные (более 75-80%о). Увеличение солёности происходит или в сильно изолированных заливах (Сиваш, Кара-Богаз гол и т. п.), или в бессточных котловинах. В обоих случаях оно происходит из-за превышения испарения над притоком. При сохранении связи с морем гипергалинные воды населяют преимущественно эвригалинные виды морского происхождения. Ультрагалинные и гипергалинные водоёмы, не связанные с морем, населяют эвригалинные виды явно пресноводного происхождения (водоросль Dunaliella sa- lina, коловратка Branchionus plicatilis, ракообразные Artemia salina, Moina mongolica, личинки комара Chironomus salinariiis и др.). Подобные биотопы изолированы друг от друга и эфемерны, поэтому населяющие их организмы образуют покоящиеся стадии, способные к дальнему разносу ветром.