Причины и скорость образования новых видов и экосистем
Итак, моделирование показывает, что для возникновения дискретности достаточно существования биотических взаимоотношений. Детальное изучение биологии отдельных видов всегда выявляет многочисленные и существенные примеры взаимодействия изучаемого вида с другими, обитающими вместе с ним.
Кажется совершенно невероятным, что все эти взаимодействия никак не влияют на эволюцию длительно сосуществующих видов. На признании этого положения и построена парадигма структурализма. Согласно этой теории в геологическом, эволюционном масштабе времени живущие совместно виды в процессе эволюции адаптируются друг к другу, образуя целостные комплексы, которые мы пока будем называть экосистемами.«Чем выше таксономический ранг, тем более длительный промежуток требуется для его появления» (Зенкевич, 1971, цит. по 1977: 227).
«Обособление таксонов разного ранга... требует тем больше времени, чем выше ранг таксона» (Беляев, 1977:211).
Однако процессы эволюции традиционно не рассматривают в экологических исследованиях, поскольку очень широко распространено мнение об эволюции как медленном, плавном процессе накопления морфологических изменений. Имеется в той или иной степени бессознательная тенденция связывать степень морфологических отличий со временем, за которое они происходят: род считают более древним, нежели вид, семейство — более древним, нежели род и т. д. В качестве примера такой точки зрения можно привести таблицу 4.2.
Уровень
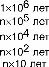
минуты - годы
Время
Семейство
Род
Вид
Морфологические изменения в популяции Функциональные изменения в популяции Индивидуальная адаптация
Цифры, приводимые в подобных таблицах, сильно зависят от анализируемых данных. Так, средний возраст современного вида насекомых — не менее 107 поколений (Же- рихин, Расницын, 1980), т.
е. соответствует возрасту семейства в приведённой таблице. Известны многочисленные примеры длительного существования видов в неизменном состоянии (латимерия, гаттерия и т. д.). Хрестоматией пример щитня Triops canacriformis, существующего до наших дней с позднего триаса, т. е. около 180 млн. лет. Есть и противоположные примеры.После вселения из Северной Америки для борьбы с амброзией амброзиевого листоеда Zygogramma suturalis в течение 5-7 поколений образовался новый подвид Z. suturalis volatus Kovalev, 2002, отличающийся как морфологией, так и способностью к полёту, которой лишены оба американских подвида (Ковалев, 2004).
Пять эндемичных видов гавайских бабочек Hedylepta встречаются исключительно на бананах, которые завезли полинезийцы на Гавайские острова около 1000 лет назад (Zimmermann, 1960).
Пять видов рыб родаHaplochromis, эндемичных для африканского оз. Набугабо, образовались не ранее 3720 лет назад (Greenwood, 1965).
Число примеров можно значительно увеличить, но совершенно ясно, что имеется весьма существенный (не менее 6 порядков) разброс скорости видообразования. Поэтому в целом можно утверждать, что представление о чёткой функциональной связи между рангом таксона и временем, за которое этот ранг был достигнут (настолько чёткой, что можно определить возраст таксонов и время изоляции биот по рангу таксонов), фактические данные не только не подтверждают, но, наоборот, позволяют однозначно утверждать, что это представление совершенно неверно.
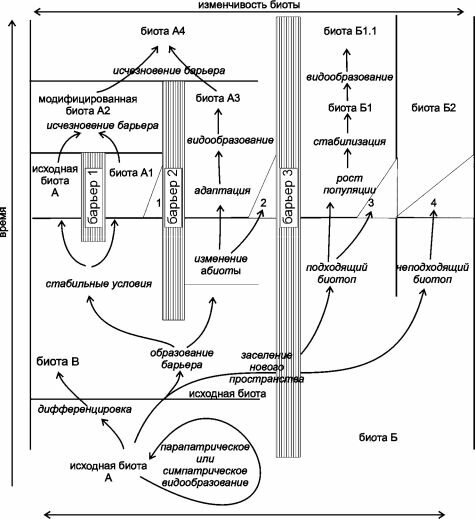
Также весьма широко распространено представление об изоляции как ведущем факторе эволюции. Классическая схема того, как возникают виды и экосистемы приведена на рис. 4.10. Согласно этим представлениям, новые виды и экосистемы образуются преимущественно после разделения единого ареала (вида, экосистемы) непреодолимым барьером. Под барьером следует понимать любую преграду, исключающую обмен генами между популяциями по разные стороны барьера. На схеме исходной биотой является биота А. Возникшие барьеры (барьеры 1 и 2), разделяют ареал этой биоты, в каждом из ареалов возникают самостоятельные биоты, которые постепенно дивергируют.
В случае исчезновения барьера (барьер 1), биоты сливаются в одну. Аналогично развиваются события после проникновения биоты через обычно непроницаемый для неё барьер (барьер 3). Выдающийся американский эволюционист Эрнст Майр (1905-2005) вообще отрицал возможность симпатрического (без изоляции) видообразования, за исключением особых случаев, вроде полиплоидного видообразования. Поскольку на его трудах (Майр, 1947, 1968, 1971, 1974) выросло не одно поколение биологов, как за рубежом, так и у нас в стране (да и я в том числе), представление о главенствующей роли изоляции в видообразовании получило широчайшее распространение, и его считают одной из прописных истин.Тем не менее, изучение реальных событий показывает, что представление о необходимости изоляции для образования новых видов, биот и экосистем отнюдь не всегда, а, возможно, редко соответствуют действительности.
Уже беглый взгляд на распределение видов по Земле показывает, что разные районы населены разным набором видов. Несомненно, что одни виды произошли от других, но
пространство
Рис. 4.10. Распространённое представление об образовании видов и биот. По Myers, Giller (1988). Объяснения в тексте .
при этом часто совершенно невозможно представить себе физические преграды, разделившие гипотетический ареал предкового вида, особенно сложно это сделать в случае морских видов, особенно учитывая их широчайшие возможности для распространения благодаря долгоживущим пелагическим личинкам, которые имеет подавляющее большинство видов бентоса. Так, одни виды образовались в высоких широтах, после произошедшего в олигоцене - миоцене увеличения температурного градиента между экватором и полюсом: в конце палеоцена поверхностная температура воды в Антарктике достигала 18°С. Другие виды сформировались в приэкваториальных районах, после произошедшего в то же время повышения температуры выше экологической амплитуды существовавших ранее в этих местах экосистем.
Третьи виды характерны для больших глубин Мирового океана. Ни в одном из этих примеров (а число их можно значительно увеличить: это и ультраабиссальные желоба посреди абиссальных равнин, и глубоководные котловины посреди шельфа (Ледовитый океан, Японское море), и некоторые крупные озёра (Байкал, Ньяса и т. п.) не возникала преграда, ареал новых видов всегда оставался связанным с ареалом предковых, их ареалы в некоторых случаях граничили на протяжении тысяч километров! И, если в отношении тропических лесов имеются данные о том, что в некоторые периоды их площадь резко сокращалась и сплошные массивы распадались на отдельные части, то современная фрагментация бореальных районов моря близка к максимальной за всё время их существования, а представить барьер, отделяющий малые глубины от больших, ультраабиссальные желоба от абиссальных равнин, вершины гор от их подножий не стоит и пытаться.«Опишите процесс видообразования у видов с облигатным бесполым размножением» (Вопрос экзаменатора «на засыпку»).
«Close relatives (sister species) have initially vicariant ranges, sympatry is secondary, resulting from later dispersal» (Banarescu, 1992: 823). Э. Майр написал предисловие к этой монографии.
«Для образования эндемичных видов требуется не только географическая изоляция, но и продолжительное время» (Абдурахманов и др., 2001: 355).
«Наличие физических преград, хоть в какой-то мере ограничивающих обмен биотой между территориями со специфическими условиями жизни, становится обязательным условием формирования региональной флоры и фауны» (Мордкович, 2005: 138).
Изучение геологической истории показывает, что генезис путём изоляции, хотя и имеет место, но встречается редко. Образование видов в результате возникновения барьеров, разделяющих исходную популяцию на две разные — способ гораздо менее распространённый, чем это принято считать. Он просто удобен с педагогической точки зрения для объяснения эволюции и видообразования. В действительности географическая изоляция не только не является необходимой для эволюции, но даже не стимулирует её.
Конечно, изоляция может приводить к постепенной дивергенции разделённых частей ранее единой экосистемы и возникновению из неё дочерних. При этом из одного предкового вида произойдут два, по одному в каждой из таких частей. Пример этому мы имеем в превращении единой изначально биоты моря Тетис, распавшейся на ряд тепловодных биот. Другой хорошо документированный пример — сравнительно недавнее разделение единой тропической морской центральноамериканской биоты после образования Панамского перешейка примерно 3,5 млн. лет назад. В этом случае по разные стороны перешейка сформировались разные экосистемы и виды, их образующие.
Но даже в таких, казалось бы, хрестоматийных примерах видообразования в результате географической изоляции при детальном анализе оказывается не всё так очевидно. Прежде всего, нет оснований считать, что все различия обусловлены только и исключительно самим фактом изоляции. Каждая из ставших изолированными частей имеет свою дальнейшую историю, в них независимо менялись климат и структура биотопа. После разделения Панамским перешейком части ранее единой биоты взаимодействовали с разными биотами. Распространённость разных сочетаний абиотических условий в изолированных биотопах может различаться. Это приводит к изменению площади, которую могут занять разные сообщества. Следствием этого является сокращение разнообразия одних и рост разнообразия других. Эти и другие причины не могли не повлиять на их современный облик и таксономический состав в частности. Так, хотя видовой состав многих таксонов по разные стороны Панамского перешейка весьма сходен, фауны рифообразующих мелководных кораллов Восточной Пацифики и Карибского бассейна совершенно различны (см. главу 3 и рис. 3.60). Фауна кораллов Восточной Пацифики несомненно является производной фауны Западной Пацифики, а вовсе не карибской, как можно было бы ожидать, учитывая недавнее возникновение Панамского перешейка. Это свидетельствует о том, что после образования Панамского перешейка мелководные рифообразующие кораллы в Восточной Пацифике вымерли.
А ведь с коралловыми рифами связано большое число видов других групп. Поэтому вымирание кораллов неизбежно должно было привести к вымиранию видов и существенному изменению и перестройке не только экосистем самих коралловых рифов, но и рыхлых грунтов.А.М. Дьяконов (1955) указывает на наличие тропических видов, обитающих как у тихоокеанского побережья Америки, так и в Индо-Малайском архипелаге, но отсутствующих в Вест-Индии, скорее всего причина этого — расселение видов после образования Панамского перешейка.
К.Н. Несис (1985), сравнивая тропические фауны головоногих моллюсков по обе стороны Панамского перешейка, отмечает, что число бентосных и океанических видов в обоих районах примерно одинаково, тогда как нектобентосных и неритоокеанических гораздо меньше в восточнотихоокеанском районе (5 против 28). Он объясняет это резким интенсифицированием и слиянием Перуанского и Калифорнийского апвеллингов в плейстоцене, приведшим к уничтожению биотопа тропической нижнесублиторальной биоты (внешний шельф - верхняя часть склона, очень узкий в этом районе) из-за постоянного снижения температуры и концентрации кислорода, характерного для апвеллингов.
Луга морских трав весьма обычны в Карибском районе, но отсутствуют в тропической восточной Пацифике, как и в тропической восточной Атлантике (Phillips, Menez, 1988). Поскольку морские травы известны с мела, нет сомнений, что они обитали в районе Панамского перешейка до его образования, и вымерли в тропической восточной Пацифике лишь в самое последнее время. Многие обитатели сообществ морских трав Кариб- ского бассейна имеют близкие виды по другую сторону Панамского перешейка, где они обитают в сообществах рыхлых грунтов. Всё же способности видов менять свою биологию конечны, поэтому часть видов, населявших сообщества морских трав до образования Панамского перешейка, вымерли с его тихоокеанской стороны, как и сами эти травы, а сохранившиеся виды изменились.
Нет в тропических восточных районах Пацифики (как и Атлантики и Индийского океана) и брахиопод — типа, известного с палеозоя — времени, когда ещё не было разделения Мирового океана на Тихий, Атлантический и Индийский (Зезина, 1973).
С другой стороны, число вымерших в Вест-Индии родов моллюсков больше, чем в Восточной Пацифике (Vermeji, 1978), что свидетельствует о том, что и в Вест-Индии условия существования морской биоты претерпевали значительные изменения. Факт массового вымирания тропической фауны Западной Атлантики в это время показан для моллюсков (Stanley, Campbell, 1981).
Итак, таксоцены неритоокеанических и нектобентосных головоногих моллюсков, иглокожих и брахиопод и сообщества морских трав и коралловых рифов (совершенно между собой не связанные) показывают, что после возникновения Панамского перешейка в тропиках восточной части Тихого океана и западной Атлантики произошли существенные климатические изменения, приведшие к крупномасштабным вымираниям. И климатические колебания, и крупномасштабные вымирания не могли не оказать влияния на эволюцию сохранившихся видов.
G.J. Vermeji (1978), сравнивая тропические восточнотихоокеанскую и западноатлантическую биоты, обнаружил, что степень морфологического расхождения гастропод, декапод и стоматопод по разные стороны Панамского перешейка ниже у видов, населяющих рыхлые грунты мелководий, нежели у видов, населяющих каменистые грунты. Все виды тропических американских двустворчатых моллюсков с большим зиянием сзади (т. е. глубоко закапывающихся) имеют пары близких видов по обе стороны Панамского перешейка[********]. Этот и другие разобранные G.J. Vermeji примеры привели его к заключению, что скорость расхождения таксонов после разделения единого ареала выше в более гетерогенных биотопах.
Поэтому таксономическое различие биот по разные стороны Панамского перешейка лишь частично можно объяснить самим фактом их географической изоляции. Очевид-
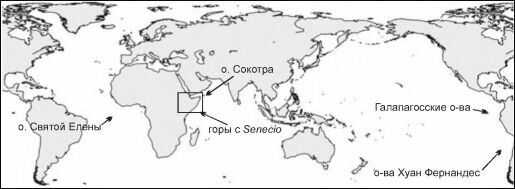
Рис. 4.11. Районы с дефицитом деревьев.
но, что и в других случаях роль географической изоляции трудно отделить от роли различной истории участков, ставших изолированными.
В то же время имеются многочисленные примеры того, что виды образовывались без изоляции.
Рассмотрим некоторые примеры районов, в которых образовывались специфичные для них виды (4.11), а затем попытаемся понять, почему в этих районах шёл интенсивный процесс видообразования, тогда как в других — нет.
Еще по теме Причины и скорость образования новых видов и экосистем:
- Эволюция островных экосистем. Динамика видового состава экосистем
- ПРИЧИНЫ ОБРАЗОВАНИЯ БОЛОТ И ИХОСОБЕННОСТИ
- Г лава 5 РАЗМЕЩЕНИЕ И ПЕРЕМЕЩЕНИЕ НАСЕКОМЫХ НА ТЕРРИТОРИИ. ФОРМИРОВАНИЕ НОВЫХ РАС И ВИДОВ
- 14* Образование видов и остальных таксонов
- Изменения экосистем. Сопряженная эволюция разных видов (коэволюция)
- Образование льда Вертикальная циркуляция и образование льда в пресной воде
- 1.3.3. Видовой состав возбудителей Malassezia-инфекций и их распространенность у животных различных видов
- Скорость почвообразования при вмешательстве человека
- 9-1. Рост разнообразия и скоростей
- Факторы, влияющие на скорость резорбции через кожу.
- Эффективность и скорость действия естественного отбора
- О СКОРОСТИ ОБНОВЛЕНИЯ БЕЛКА И ХЛОРОФИЛЛА В ВЫСШИХ РАСТЕНИЯХ [24]
- Механизм распространения и скорость световых волн
- Ведущая роль отбора в возникновении новых признаков