Двустворчатые моллюски Арала
На Аральском море человек в конце прошлого века поставил крупномасштабный эксперимент по образованию новой экосистемы. С резким сокращением, а временами и полным прекращением пресного стока в море, произошло стремительное нарастание солёности и общее уменьшение площади водоёма (рис.
4.18). До зарегулирования стока рек основными первичными продуцентами являлись макрофиты (Карпевич, 1975), но к 1990- м гг. большинство видов макрофитов вымерло: из 37 сохранилось только 4 (Доброхотова, 1971; Orlova et al., 1998). Это привело к резкому возрастанию содержания биогенов в воде, несмотря на прекращение их поступления с речным стоком. В результате, к началу 1990-х гг. произошло резкое возрастание концентрации фитопланктона, и он стал, как и в остальных морях, основным продуцентом. Произошедшие изменения биотопа и продукционного блока вызвали соответствующие перестройки трофической структуры и таксономического состава зоопланктона, зообентоса и ихтиофауны: из 49 видов зоопланктона сохранилось только 7, из 61 видов зообентоса — 27, из 33 видов рыб — 9 (Андреева, 1989; Андреев, 1999). Фактически за период менее 30 лет сформировалась совершенно новая экосистема. Детально были проанализированы изменения, произошедшие с Bivalvia (Андреева, 2001; Андреева, Андреев, 2001, 2003). Массовое вымирание привело к появлению и накоплению в Арале неиспользуемых ресурсов: к первой половине 1980-х гг. уцелел случайный набор видов, некоторые трофические группировки были полностью утрачены. В зоопланктоне, зообентосе и нектоне исчезли практически все хищники. В зообентосе сохранились только собирающие детритофаги и один фильтратор инфауны. Несмотря на резкое увеличение концентрации фитопланктона, потреблявшие его фильтраторы эпифауны вымерли. Для потребления неиспользуемых ресурсов симпатрически возникли новые таксоны, в первую очередь из предкового вида Cerastoderma glaucum (= С. umbonatum, С. lamarcki, С. isthmicum), перешедшего от фильтрации из прилежащего к грунту слоя воды (фильтратор инфауны) — тип питания большинства Cardiidae, к фильтрации из толщи воды (фильтратор эпифауны рыхлых грунтов) — тип питания, вообще не свойственный со-
Рис. 4.18. Аральское море в 2000 г., видна береговая линия середины прошлого века. По Аладину
и др., 2004).
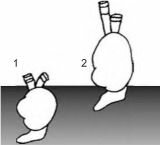
временным Cardiidae (в Паратетисе таксоны Cardiidae с таким типом питания возникали неоднократно). Освоение новой адаптивной зоны сопровождалось «рядом аро- морфных преобразований... из наиболее существенных изменений морфологии раковины следует отметить новые варианты конфигурации створок с удлинением заднего края раковины и сильным смещением вперёд макушек, уплощение раковины, резкое уменьшение числа рёбер, изменение строения лигамента и замка. В начале 1990-х гг. сформировалось несколько групп, изменчивость между некоторыми из них превышает межвидовую изменчивость современных и ископаемых Cerastoderma, Рис. 4.19. Разные экотипы а по отдельным морфологическим признакам (замок, ли-
Cerastoderma Аральского мо- гамент) выходит за пределы рода и даже семейства» (Ан-
ил и песчаный ил
тонкодисперсный ил
Abra segments Hypanis Cerastoderma glaucum
илисто-ракушечный грунт и песок
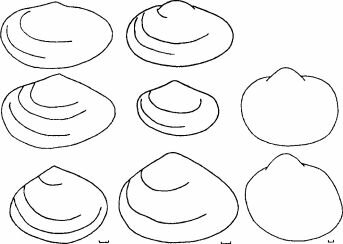
Рис. 4.20. Жизненные формы инфаунных двустворчатых моллюсков Арала. По С.И. Андреевой (2001).
ря. По С.И. Андреевой (2001). дреев, Андреева, 2001: 42) (рис. 4.19). Материнская форма также сохранилась.
В другой из переживших перестройку групп Bivalvia — АЪга — из одного материнского вида П. segmenta (= Syndosmia segmentum), также недавнего вселен- ца из Средиземного моря, образовалось три формы. Эти формы поразительно соответствуют видам Hypanis, вымершим в Арале в результате перестройки (рис. 4.20). В обоих случаях (с Cerastoderma и с АЪга) новые формы длительно и устойчиво сосуществуют, что позволяет говорить об их таксономической самостоятельности. Таким образом, за 10-15 поколений образовалось несколько видов моллюсков, с разным типом питания и столь сильно различающихся морфологически, что их (в случае с Cerastoderma) можно отнести, как минимум, к разным родам, а через пару десятков лет, при сохранении той же скорости эволюции, возможно, и к разным семействам. Ни о каком аллопатрическом видообразовании не может идти и речи: все возникшие за это десятилетие виды симпатрич- ны. Нет оснований предполагать наличие каких-либо прекопуляционных механизмов изоляции: все они имеют наружное оплодотворение и пелагическую личинку.Абиссальная фауна Средиземного моря
Средиземное море (площадь — 2510 млн. км2, объём — 3771 млн. куб. км) соединено с Атлантическим океаном узким Гибралтарским проливом глубиной 350 м, и разделено на две части Сицилийским проливом глубиной 400 м. Максимальная глубина восточной части — 5093 м, западной — 3730 м. В настоящее время менее плотная атлантическая вода поступает в Средиземное море по поверхности, тогда как глубины заняты более плотной и солёной водой местного происхождения, она изливается в придонных слоях через Гибралтарский пролив в Атлантику. Температура воды в самых глубоких частях Средиземного моря гораздо выше, чем где бы то ни было в Мировом океане на таких глубинах — более 13°С (в Красном море температура на глубинах также высока, но на абиссальных глубинах там нет макрозообентоса, они заполнены бескислородным рассолом). В результате климатических колебаний начиная с конца миоцена (подробнее — см. раздел «История Мирового океана») в Средиземном море периодически устанавливалась вертикальная стратификация, приводящая к появлению сероводорода на больших глубинах моря, по типу современного Чёрного моря.
Последний такой период был между 25000- 23000 и 9000-6800 лет назад. В это время в восточной части моря образовался слой сапропеля толщиной 2,5-70 см (Buckley, Johnson, 1988). Во время этих периодов глубоководная биота, по-видимому, полностью вымирала. Тем не менее, в глубоководной части Средиземного известно 20-30 эндемичных ви- дов, принадлежащих частично к родам, обитающим и на малых глубинах, а частично — к глубоководным родам (Пастернак, 1982; Bianchi, Morri, 2000). Формирование эндемичной глубоководной фауны, пускай и немногочисленной, показывает, что даже на больших глубинах скорость видообразования может быть весьма высокой.Крупные озёра
Байкал — классический в отечественной биогеографической литературе пример региона с высоким эндемизмом. Причины этого эндемизма и обособленности байкальской фауны разные авторы объясняют, казалось бы, по-разному. Одни (например, Верещагин, 1940) объясняют отличия байкальской фауны, её обособленность, происхождением от фауны мезозойских морских бассейнов Азии. Другие, (например, Берг, 1948) считают, что обособленность фауны Байкала от окружающей связана с вымиранием последней, т. е. считают байкальскую фауну реликтовой. Общим для этих точек зрения является то, что их сторонники считают, что фауна Байкала богата эндемиками потому, что она является реликтом какой-то ещё более богатой фауны, т. е. сам Байкал рассматривают как рефуги- ум. Поэтому их можно объединить в одну гипотезу реликтов.
Эта гипотеза в действительности объясняет отнюдь не все факты; остаётся неясным:
«Предполагается, что причиной несмешиваемости фаун Байкала и окружающих водоёмов служит особое строение байкальской воды, содержащей примесь изотопических молекул, увеличивающих её плотность» (Бирштейн, 1946: 244).
- почему существующие отличия Байкала и окружающих водоёмов столь резки, т. е. почему реликты сохранились почти исключительно в Байкале (а также водоёмах с условиями, сходными с байкальскими, в которые они могли попасть из Байкала)? Для объяснения этого выдвигали совсем уж фантастические теории.
- почему высокий эндемизм обеспечен на основе очень небольшого числа предковых форм (табл. 4.3)?
Виктория
Верхнее
Танганьика
Ньяса
Таблица 4.4. Морфометрические данные некоторых крупных озёр.
озеро площадь глубина, м
тыс. км2 максимальная аэробной зоны
Верхнее 82,4 393 393
Виктория 68 80 80
Танганьика 34 1470 70-240
Байкал 31,5 1620 1620
Ньяса 30,8 706 35-50
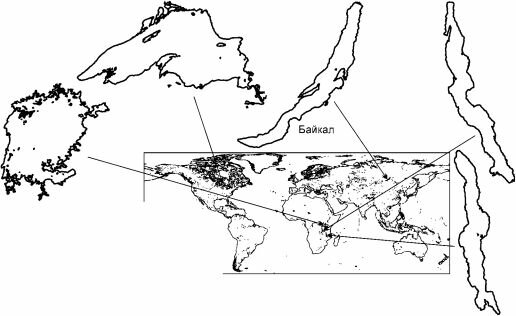
Рис. 4.21. Некоторые крупные озёра в одном масштабе.
Если рассматривать проблему шире, не замыкаясь на Байкале, а анализируя сходные биоты, то оказывается, что обе эти особенности свойственны и другим крупным озёрам с высоким эндемизмом, но совершенно другой историей. В отношении этих озёр также выдвигали теории о происхождении их биоты из морской (примеры см.: Старобогатов, 1970). Объяснить озёрный эндемизм морским происхождением пытались даже для Иссык-Куля, расположенного на высоте 1608 м над уровнем моря! Одни из таких озёр с высоким эндемизмом, как и Байкал,
Таблица 4.3. Число предполагаемых исходных видов и число развившихся из них таксонов в фауне Байкала. По Кожову (1962), Семерному (2004), Ка- малтинов (2002); Камалтинов (2002); Strong et al. (2008).
таксон число эндемичных
исходных видов родов семейств видов (подсемейств)
| губки | 1-2 | 6 | 3 | 1 |
| турбеллярии | 3-4 | до 40 | ? | - |
| олигохеты | ? | 164 | 11 | - |
| остракоды | 1-2 | 19 | - | - |
| изоподы (Asellus) | 2 | 5 | ||
| Gammaridae | 4-5 | 363 | 72 | 7 |
| ручейники | 2-3 | 10-12 | 2 | |
| гастроподы | 5-6 | 114 | 6 | 3 |
| рыбы | 2-3 | 23 | 7 | 3 |
представляют собой трещины в коре, другие имеют большую площадь, но сравнительно мелководны (табл. 4.4, рис.
4.21). В качестве примера можно привести озеро Ньяса (Малави). Это озеро возникло примерно 500 тыс. лет назад (современный глубоководный режим установился там гораздо позднее — примерно 50 тыс. лет назад), из примерно 250 видов обитающих тут рыб, более 90% — эндемы: около 100 эндемичных видов широко распространённого рода цихлид Haplochromis, 70 других видов цихлид, принадлежащих к 20 эндемичным родам (Banarescu, 1995) (рис. 4.22) и несколько эндемичных видов рыб других семейств, по другим данным в нём обитает более 200 видов цихлид, из которых —
Petrotilapia tridentiger оброет камней

Аи ion оса га nyassae собирает членистоногих
Labidichromis
vellicans
собирает
членистоногих
Genyochromis
mento
рыбы
Docimodus
johnstoni
рыбы
Haplochromis pardalis рыбы
Haplochromis polyodon рыбы
Cyathochromis obliquidens оброет камней и листьев

Hemitilapia
oxyrchynchus
оброет
растений


Labeotropheus (5) fuellebornii оброет камней
Corematodus
chiranus
рыбы
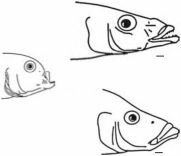
Haplochromis compressiceps — рыбы
lies (1972). Указан объект питания.
Rhamphochromis macrophthalmus рыбы
Pseudotropheus
fuscus
оброет камней

Lethrinops brevis насекомые, зарывшиеся в песок

см. По Fryer,
Рис. 4.22. Цихлиды оз. Ньяса. Все шкалы — 1
Haplochromis . , fenestratus ъ (V водоросли, неспециализированный
Haplochromis
guentheri
нитчатые
водоросли

Pseudotropheus tropheops оброет камней

Haplochromis
similis
листья
Pseudotropheus zebra оброет камней

Haplochromis euchilus насекомые на камнях
эндемичны 180 (Махлин, 1983) или 178 видов, из которых 174 — эндемы (Дарлингтон, 1966). Эндемичные цихлиды образовались из двух исходных видов (Roberts, 1975). Впадина озера Танганьика в три раза старше (1,5-2 млн. лет), но в последние 50-60 тыс. лет уровень озера неоднократно снижался на сотни метров (до 600 м). Специфичность ихтиофауны озера примерно та же на видовом уровне и выше на родовом: из общего числа примерно 200 видов (примерно 40 родов) 95% — эндемы. Несомненно, что условия и имеющие ресурсы в этих огромных озёрах существенно отличны от таковых в окружающих мелких водоёмах и реках. Поэтому в этих озёрах не могли функционировать экосистемы окружающих водоёмов. В полном соответствии с теорией структурализма при образовании значительной территории, не могущей быть населённой ни одной из существующих экосистем, в них сформировались самостоятельные экосистемы, о чём и свидетельствует образование эндемичных для них видов. Принять участие в формировании новых биот в силу разных причин смогли немногие виды, именно поэтому многочисленные эндемичные виды образовались из немногих исходных, как, например, в Байкале (табл. 4.3). Новые виды образовывались для потребления специфических для данного озера ресурсов, в разных озёрах эти ресурсы разные, различны и донорские биоты, поэтому в каждом озере в одних группах наблюдается высокое разнообразие, тогда как другие или малоразнообразны, или вовсе отсутствуют. Группы, дающие вспышки видообразования в разных озёрах системах, различны. Так, в частности для тропических африканских озёр характерен высокий эндемизм рыб и наяд (группы двустворчатых моллюсков, куда входят беззубки и перловицы), тогда как фауна бокоплавов и изопод (таксонов разонообразие которых в тропиках гораздо ниже, чем в умеренных широтах) крайне бедна, в Байкале же, расположенном в умеренных широтах, исключительно велико разнообразие бокоплавов и изопод, эндемичных рыб немного, а эндемичных наяд в нём вовсе нет.
Скорость образования видов (эндемов) при формировании новой экосистемы поис- тине сказочно велика, что находится в резком противоречии с общепринятыми представлениями о скоростях видообразования и вообще об эволюции, как плавном и неторопливом процессе. Так, согласно современным данным, оз. Виктория полностью высохло в позднем плейстоцене, примерно 12 тысяч лет назад (Johnson et al., 1996). Очевидно, что как минимум булыпая часть видов биоты существовавшего до того озера вымерла. Тем не менее, современная биота озера насчитывает только цихлид более 300 эндемичных видов (Brown, Lomolino, 1998). По другим данным (Banarescu, 1995) в озере около 200 видов, из которых 169 — эндемы (Махлин, 1983: 160-170 видов цихлид, из них лишь три — неэндемичные). Очевидно, что для образования бульшей части эндемичных видов оказалось достаточно этих 12 тысяч лет. Очевидно также, что видообразование происходило несомненно симпатрически без какой-либо изоляции, для происхождения видов аллопатриче- ским путём не было ни условий, ни времени.
Озеро Ланао (Lanao) на Филиппинах почти в 200 раз меньше озера Виктория (375 км2) и имеет примерно тот же возраст (10 тыс. лет). Неїте в 1924 и 1932 гг. описал 13 эдемичных видов (но не цихлид, а карповых Cyprinidae), относящихся к 1 широко распространённому и 4 эндемичным родам, три из которых монотипичны, а один включает два вида. Эти виды несомненно произошли также от одного современного вида Pimtius Ыпо- tatus (= Barbus binotatus), широко распространённого на Филиппинах. При этом у Mandi- bularca и Spratellicypris строение нижней челюсти выходит за пределы вариабельности во

всем семействе карповых (275 родов и более 1700 видов). Их валидность одними была признана (Myers, 1960), другими — нет (Banarescu, 1992). Основной аргумент P. Banares- cu, в принципе отрицающего возможность симпатрического видообразования: «evidently not consistent with princeples of the strictly phyletical (hennigian) classification». Также он указывает, что три из пяти видов эндемичных родов были описаны по одному экземпляру, а голотипы утрачены во время войны. В дальнейшем, несмотря на интенсивные попытки, не удалось их поймать, правда озеро с тех пор подверглось сильному антропогенному воздействию и они могли вымереть (Leveque et al., 2008). В то же время P. Banarescu признаёт, что краб Archipelothelphusa wolterecki действительно эндемичен для Ланао. Относительно малакофауны озера Я.И. Старобогатов отмечает, что «почти все виды, отмеченные здесь, эндемичны для озера» (1970: 210).
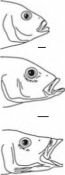
У обитающих в Танганьике цихлид строение зубов также выходит за пределы вариабельности, но даже не семейства, а всего класса: у видов эндемичного Perissodus зубы двувершинные или листовидные (рис. 4.23), приспособленные для скусывания чешуи рыб — основной пищи взрослых рыб этого рода. Характеризуя это явление G.S. Mayers и ввёл понятие «запредельная специализация».
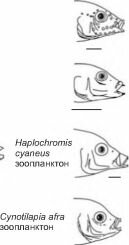
Известны многочисленные примеры быстрого образования рас рыб в крупных водоёмах для освоения новых ресурсов (отсутствовавших ранее или потреблявшихся другими, специализированными ранее видами). В наиболее распространённом случае из планкто- фага или эврифага образуется две морфы: крупная хищная и мелкая, обычно более тёмно окрашенная, питающаяся планктоном.
Балтийское море. Салака Clupea harengus membrans существует в двух формах: типичной и гигантской (гигантские стремлинги). Типичная салака питается преимущественно мелким зоопланктоном, реже зообентосом и в возрасте 5-6 лет достигает длины 16-20 см. Стремлинги питаются преимущественно рыбой (в том числе и салакой). Длина стремлингов в 1,5-2 раза превышает длину типичной салаки того же возраста (Светови- дов, 1952). Время вселения салаки в Балтийское море — примерно 4000 лет назад (после осолонення Анцилового моря-озера).
Байкал. Эндемичные для Байкала голомянки (Comephoridae) включают два пелагических вида: малую (Comephorus dybowskii) и большую (С. baicalensis) голомянку. Малая — планктофаг, большая — хищник.
Дополнительные примеры можно найти у Б.М. Медникова (1963, 2005), М. Barluen- ga et al. (2006).
Таким образом, одновременно с образованием крупных озёр (или других крупных водоёмов), условия в которых резко отличаются от условий в окружающих водоёмах, в них формируются экосистемы из имеющихся в данной местности. В результате некоторые таксоны преобразуются в эндемичные. К аналогичным выводам пришёл и Я.И. Старобогатов (1970), анализируя пути формирования фауны моллюсков озера Танганьика. Время, необходимое для формирования новой экосистемы озера, в геологическом масштабе может быть ничтожно малым, и небольшим даже по человеческим меркам. Из этого, в частности, следует то, что эндемичная фауна Байкала автохтонная, образовалась она в самом Байкале, а большой возраст её видов, возможно, сильно преувеличен.
Еще по теме Двустворчатые моллюски Арала:
- КЛАСС ПЛАСТИНЧАТОЖАБЕРНЫХ, ИЛИ ДВУСТВОРЧАТЫ
- Другие двустворчатые и их промысловое значение
- ПО СТОПАМ МОЛЛЮСКОВ
- Моллюски
- Моллюски
- Моллюски
- Моллюски
- Брюхоногие моллюски
- ГОЛОВА — НЕ САМОЕ ВАЖНОЕ
- Проверка двух теорий эволюции
- 2. Функционирование биоценозов
- ФИЛЬТРАТОРЫ
- Формы акклиматизации.
- Дрейссена и ее особенности
- Горчак (Rhodeus cericeus)
- РАЗРУШИТЕЛИ КОРАЛЛОВЫХ РИФОВ
- ВЕТВИСТОУСЫЕ И ВЕСЛОНОГИЕ.
- 2-6. Знаменитый спор
- Малакология