Математическое моделирование динамики популяций
В последние десятилетия специалисты по динамике популяций, как отмечалось выше, анализируют в первую очередь изменения численности особей (Воронов, 2005). Это наиболее видимый и просто регистрируемый показатель состояния популяции, но по своему характеру динамика численности особей в популяции - параметр вторичный, он является результатом предварительного изменения состояния особей, а также онтогенетической и виталитетной структуры популяции.
Тем не менее основные математические модели популяций ориентированы на анализ динамики численности особей.Основной механизм, лежащий в основе динамики популяций с точки зрения изменения численности особей в них - это соотношение рождаемости и смертности особей. Такие модели строятся на основе самых общих представлений о динамических процессах и способствуют выявлению и оценке значимости факторов, лежащих в основе динамики популяций. Некоторые из моделей представляют собой компьютерные программы, но как правило, многие математические модели непригодны для реальной оценки динамики природных популяций, так как, с одной стороны, они не учитывают ряда важных факторов, действующих на популяции в конкретных условиях, а с другой - требуют знания удельной скорости роста численности особей, размеров смертности и рождаемости, а также других параметров, точная информация о которых у исследователя обычно отсутствует. Игнорируют такие модели и «неопределенности* М. Шаффера. Поэтому после некоторого бума математического моделирования динамики популяций интерес исследователей к нему заметно упал.
В самом общем виде динамические процессы в популяциях обычно представляют уравнением, которое включает в себя только один аспект этого комплексного явления — изменение числа особей в популяции:
Nt + l = Nt+B-D + I-E,
где Nt - численность особей популяции в исходный момент времени;
w, + 1 ” численность особей спустя некоторый период времени;
В - прибавка количества особей в популяции за счет рождаемости;
D - убыль особей за счет смертности;
I - иммиграция особей;
Е - эмиграция особей.
Интервал t... t + 1 должен соответствовать биологии организма, то есть скорости размножения и отмирания. Для растений иммиграция и эмиграция происходят только в ходе распространения семян и, следовательно, имеют сдвиг (лаг) во времени.
При равновесии процессов размножения и иммиграции, с одной стороны, и процессов смертности и эмиграции - с другой, популяции находятся в стабильном состоянии. Превышение процессов смертности и эмиграции особей ведет к снижению популяционной плотности, инсуляризации и деградации популяции, тогда как превышение процессов рождаемости и иммиграции имеет следствием рост популяции.
Естественно, что размножение и смерть в мире растений не могут быть постоянными величинами, и поэтому правильнее эти параметры выражать как удельные, то есть отнесенные к одной особи;
. Bjt). t N(t)’
At N(t)
Тогда рождаемость и смертность можно записать так:
B{t) = bN(t)At и D{t) = dN{t)At.
Подставив полученные значения в исходную формулу и выполнив необходимые преобразования, получаем основную формулу для модели экспоненциального роста популяции;
42L=rN(t).
dt
Экспоненциальная кривая роста, или, как ее еще называют, J-об- разная кривая, была изучена и описана А. Лотокой и В. Вольтерра
(рис. 9.1а). Такие кривые роста наблюдаются в природных условиях не так часто, но и не редко. Экспоненциальный тип роста популяции может наблюдаться в полностью свободных экологических нишах и только на протяжении короткого отрезка времени. Он также характерен для популяций инвазионных видов растений, которые заселяют новые для них местообитания, не имеют в них ограничений в ресурсах к не подвергаются воздействию фитофагов и различных заболеваний, «/-образная кривая роста характерна для некоторых популяций в экосистемах с малой устойчивостью, то есть с бедным видовым составом, например, для сорных видов в условиях агроценозов. Такая же независимая от плотности динамика характерна и для многих пионерных видов с г-стратегией, заселяющих еще необжитые пространства или успешно вытесняющие более слабых конкурентов из их экологических ниш.
Экспоненциальный рост имеет место при «цветении* водоемов, когда в них массово размножаются зеленые или синезеле- ные водоросли. Во всех этих случаях наблюдаются как бы популяционные взрывы (в человеческом обществе их называют демографическими взрывами) - периоды быстрого роста численности популяции.
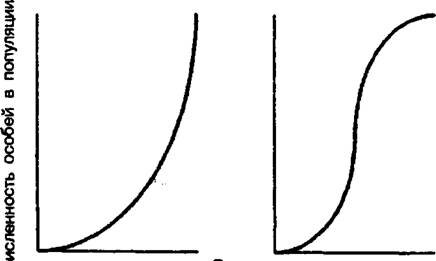
Рис. 9.1. Два основных типа кривых роста популяций:
а — экспоненциальный рост, J—образная кривая роста, б — логистический
рост, 5 - образная кривая роста
Время
х
а)
б)
Вспышки массового размножения имеют два аспекта:
- увеличивается количество растений и запас фитомассы в уже существующих популяциях;
- появляются новые популяции данного вида растения.
Иногда эти процессы протекают одновременно. Так, активная агрессия амброзии полынолистнойAmbrosia artemisifolia в Украину и в европейскую и азиатскую части России сопровождается как ростом численности существующих популяций, так и активным формированием новых популяций на территориях, где раньше амброзии не было. При этом в популяциях складывается устойчивая онтогенетическая структура с достаточно постоянным соотношением между числом особей, находящихся в разных онтогенетических состояниях.
Более адекватна динамике большинства природных популяций растений модель логистического роста, предложенная П. Фер- хюлъстом (рис. 9.16):
йК=г mK~N)
dt “ 1 к }’
где К - верхняя асимптота, соответствующая верхнему пределу численности популяции после того, как в ней наступит равновесие (ее величина определяется предельной емкостью среды). Максимально возможное значение коэффициента роста популяции гтлх, достигаемое при наиболее благоприятных условиях среды, называется биотическим, или репродуктивным потенциалом популяции.
Обычно биотический потенциал тем выше, чем ниже уровень организации организмов.Решение приведенного выше дифференциального уравнения имеет вид:
N = K/(l+eart),
где а - постоянная интегрирования, определяющая положение кривой относительно начала координат. Кривая роста популяции, описываемая данным уравнением, называется логистической, или S-образной кривой.
В рассматриваемом случае скорость роста равна максимальной скорости роста (гЫ), определяемой биотическим потенциалом, умноженной на степень реализации максимальной скорости (1 - N/К), которая тем ниже, чем больше плотность (или численность) N популяции. Поэтому такую динамику называют зависимой от плотности (численности), в отличие от независимой от плотности экспоненциальной динамики.
В принципе J-образную кривую можно рассматривать как неполную S-образную кривую, просто лимитирующие факторы среды в этом случае ограничивают рождаемость еще до того, как существенную роль в регулировке численности начнут играть внутренние факторы, обеспечивающие устойчивость системы.
Всем растениям свойственна тенденция к размножению, и поэтому в оптимальных условиях популяции расширяются и количество особей в них увеличивается. По Р. Уиттекеру (1980) в обоих этих процессах можно выделять две последовательные фазы: Я-фазу, когда численность особей возрастает по экспоненте, и Я-фазу, когда рост численности становится асимптотическим и рост популяции начинает соответствовать S-образной кривой.
При использовании математических моделей роста популяций, а их, кроме рассмотренных, современными исследователями предложено достаточно много, следует иметь в виду, что все они исходят из серьезных упрощений закономерностей популяционной жизни растений. В них допускается, что все особи в популяции одинаковы по строению и свойствам, рост популяции находится в линейной зависимости от ее начальной плотности, реакция на изменение плотности в популяции происходит мгновенно. Реально в природных популяциях ни одно из этих допущений не действует.
В целом, приходится признать, что разработка моделей динамики (да и вообще функционирования) популяций растений остается лежащей в области так называемого парадокса Бонини. В исходной формулировке П. Валери (1937) этот парадокс звучит так: всё простое неверно, а всё, что является комплексным, непригодно к использованию. Позже М. Даттон (1971) сформулировал этот парадокс следующим образом: как только модель комплексной системы становится более совершенной, она делается менее понятной. В настоящее время исследователь должен отдавать предпочтение прагматичным моделям, описывающим и прогнозирующим элементарные популяционные процессы. В первую очередь должны быть выявлены системы лимитирующих факторов, воздействующих на функционирование популяций растений.
Еще по теме Математическое моделирование динамики популяций:
- ЗАКОНОМЕРНОСТИ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ. МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ ПОПУЛЯЦИЙ
- МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ ЭМИССИИ МЕТАНА ИЗБОЛОТ В АТМОСФЕРУ
- Математические модели в генетике популяций и в теории эволюции
- Компьютерное молелирование динамики популяций
- ДИНАМИКА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
- Флюктуаиионная и сукиессионная динамика популяций
- 3. Динамика численности популяций
- Глава 9 Динамика популяций
- Динамика численности популяции во времени.
- § 1. РОЛЬ ДИНАМИКИ ВОЗРАСТНОЙ СТРУКТУРЫ ПОПУЛЯЦИИ В ПРЕОБРАЗОВАНИИ ЕЕ ГЕНЕТИЧЕСКОГО СОСТАВА
- Математическое описание отбора
- Моделирование филогенеза
- Моделирование континуальности и дискретности
- Некоторые закономерности проникновения математических методов и идей в биологию
- Часто приписывают определенные математические способности птицам...
- Математические модели в экологии. Модели биогеоценозов
- ПРЕОБРАЗОВАНИЕ ПОПУЛЯЦИЙ. ГОМЕОСТАТИЧЕСКОЕ ИЗМЕНЕНИЕ ГЕНЕТИЧЕСКОЙ СТРУКТУРЫ ПОПУЛЯЦИЙ И микроэволюция
- Границы между популяциями, иерархия популяций
- Фазовый портрет динамики численности