13.4. Механические воздействия
Устойчивость к собственной тяжести, давлению снега или эпифитов, противодействие сгибающим и «вырывающим» силам ветра и воды, толерантность к засыпанию опадающей листвой, навеваемыми песками или склоновой эрозией, а также движениям почвы под действием гравитации или образования льда — это те критерии, которые определяют возможность или невозможность существования видов или определенных жизненных форм во многих биомах (рис.
13.10).Этими вопросами занимается биомеханика. Она дает оценку эластичности и прочности структур, выясняет причины ветровала и механизмы, с помощью которых одни растения используют другие, чтобы «без лишних затрат» подниматься к свету (вьющиеся, цепляющиеся, использующие растения как опору). Закрепляемостъ почв на склонах в значительной мере определяется механическими свойствами корней и корневищ. Равным образом механически мощные системы подземных органов гарантируют, что растения после выпаса не останутся без корней. Многие растения могут сжимать свои корни и таким образом втягивать в глубину после прорастания чувствительную точку роста (см. бокс 4.5, рис. С). Большое экологическое значение имеют те силы, которые возникают при «разрядке» механического напряжения во время высы-
хания («взрывание», выбрасывание семян). Используя тургорное давление (20 бар и более), растения способны взламывать массивные структуры и протискивать свои корни в трещины грунта. Огромную механическую работу выполняют тонкие корни (защищенные на кончиках корневым чех- ликом), пронизывающие очень плотные субстраты. Жесткость (англ. toughness) листьев — существенный компонент обороны от травоядных, для которой также служат разнообразные «защитные структуры» (колючки, шипы, образование пробки). Способность к перенесению механических на-

Рис.
13.10. Примеры механических нагрузок на растения:А— лед (Eucalyptus pauciflora, 2050 м, Снежные горы, юго-восток Австралии); В — снег (верхняя граница березового леса в Северной Швеции, 700 м); С — песок (дюны в Восточной Австралии); D — щебнистый склон (Cerastium urtiflorum в Альпах); Е — опавшая листва (с проростками бука); F — бурелом и ветровал (Picea abies, после урагана Лотар 26.12.1999 г. в Шварцвальде). Следующими примерами были бы движения почвы, затаптывание при передвижении пастбищных животных, нагрузка эпифитами и лианами (см. табл. 15.1) и т.д.
грузок, развитие физических сил и упомянутых защитных структур часто обеспечивают успешное существование вида и определяют стабильность всей экосистемы. Их экологическая значимость нередко превышает таковую физиологической способности организма к приспособлениям.
- Водный режим
Наземное существование растений возможно только при эффективно действующих корнях, несжимающейся капиллярной проводящей системе (ксилема, см. 3.2.4.2), комбинации изменяющихся (устьица) и статичных (кутикула) приспособлений к защите от испарения и вакуолизированных в различной степени тургесцентных клеток (водный потенциал, см. 6.1.4.2. 6.3.2.1). При недостатке воды растение может усиливать как ее потребление, так и способы защиты от потери воды и ее последствий. Поскольку строение и функции компонентов, обеспечивающих регуляцию водного режима, были описаны выше, речь пойдет о реакциях на дефицит влаги и будет показано, как согласованно растения управляют водным потенциалом, транспортированием водных потоков и механизмами сопротивления транспортированию; при этом они сами влияют на водный режим экосистемы.
- Водный потенциал и транспирация
Водный потенциал (см. 6.1.4.2) в определенной точке растения есть результат тока воды и сопротивления ее передвижению. С термодинамической точки зрения он означает пониженную по отношению к свободной воде доступность в точке измерения, что также может быть обозначено и как напряжение, отсасывание или отрицательное давление.
Водный потенциал будет тем негативнее, чем в большей степени он падает. Пока он не перейдет критического значения (в листьях в зависимости от вида растения оно составляет от -1,5 до -2,0 МПа), говорят, что водный потенциал лишь немного превышает фактическую степень водоснабжения растения. При достаточной влажности почвы водный потенциал листьев тем ниже (негативнее), чем выше степень транспирации. Уменьшение потенциала с увеличением пропускной способности соавнимо с давлением в водопроводе, которое падает тем больше, чем шире открывают кран. Если в почве преобладает дефицит воды, даже очень незначительная транспирация (и даже полное закрытие устьиц) не может препятствовать падению водного потенциала до очень низких значений. В одинаковой степени низкий водный потенциал в листьях может появиться в зависимости от состояния снабжения водой при очень интенсивной транспирации или при почти полном ее прекращении. Без сведений о синхронном транспирационном потоке нельзя интерпретировать водный потенциал и ответить на вопрос, действительно ли растение испытывает дефицит воды (во всяком случае не при значениях более -2 МПа).Ключ к оценке снабжения растения водой — диаграмма соотношений потенциала и транспирации (рис. 13.11). Она позволяет на графике, как правило, линейных связей, используя наклонные прямые, маркирующие возрастание гидравлического сопротивления, интерпретировать его величину и определить равновесный потенциал с почвой в ранние утренние часы (англ. predawn water potential) через регрессию потенциала у нулевой точки транспирации. Пронумеровав и соединив точки на диаграмме, отмеченные хронологически в течение одного дня, можно обнаружить, что линия соединения представляет собой поучительный гистерезис (в послеполуденные часы значения возвращаются к нулевой транспирации при более низком уровне водного потенциала по тому же пути, как эти значения «шли» утром при возрастающей транспирации). Это показывает ухудшение гидравлических условий в течение дня, что связано либо с кавитацией ксилемы (закупорка воздухом, газовая эмболия), либо с сопротивлением транспортирования в корневой системе.
Водный потенциал листьев обычно ниже, чем потенциал в ксилеме осей побегов или ствола. Равновесное давление на срезе побега или черешка листа, полученное методом барокамеры (см. рис. 6.39), есть смешанная величина для всех дистальных тканей, в основном тканей листа, почему и уместно выражение «листовой» или «побетовый» водный потенциал (но не водный потенциал ксилемы). При интенсивной транспирации происходит резкий спад потенциала от оси побега через черешок листа в направлении пластинки.
Между обратным значением водного потенциала и дефицитом влагонасыщенности WSD (см. 6.3.6) существует нелинейная зависимость (кривая давления — объема), которая дает возможность выявить эластичность клеточных стенок, осмотическое давление при полном зур- горе, а также водный потенциал в точке потери тургора, поэтому такие характеристические кривые применяют для многих растений открытого грунта. Водный дефицит вообще может возникнуть из-за эластичности клеточных стенок (в «бетонных» клетках не могло бы быть дефицита влаги, так как вода их не растягивает). Эластичность клеточных стенок есть также мера того, насколько прямо (и насколько сильно) всасывающая сила атмосферы при испарении сообщается с корневыми системами почвы. Поэтому в районах с периодическими засухами возрастает число растений с очень жесткими, малоэластичными стенками клеток. Это также одна из причин распространения склерофилии в регионах с сухими периодами.
Крайне важное с экологической точки зрения явление — кавитация ксилемы. Если капилляры при сильной транспирации приходят в состояние напряжения, может наступить прорыв воздуха и образование воздушных пузырьков (газовая эмболия, см. 6.3.5), что быстро приводит к разрыву гидравлического континуума. Так как в первую очередь у деревьев при хорошей погоде кавитация является правилом (акустически ее возникновение обнаруживается в ультразвуковой области), су шествует подозрение, что кавитация не всегда неблагоприятна для деревьев (возможно, она способствует предохранению от перегрузок).
Водный потенциал в листьях вследствие этого опускается быстрее, чем в ксилеме (ограниченный континуум силы сцепления). Есть первые указания на то, что эмболия в позднее послеполуденное время может ликвидироваться точно так же быстро, как и возникнуть в полдень, причем активную роль в этом играют паренхима ксилемы и флоэма. Если удаляют флоэму, «ремонт» ксилемы явственно замедляется. Эти факты лежат в той развивающейся области исследований, в которой тонкий анализ физиологии ксилемы еще может преподнести некие сюрпризы! При этом в центре стоят не дебаты теории сцепления (которая в принципе едва ли оспорима), а изменение кавитации, ее влияние на реальное состояние напряжения в ксилеме, функция пор и древесинной паренхимы, а также то, каким образом кси-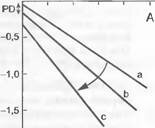
Степень транспирации, ммоль м 2 ¦ с 1 012345012345

Рис. 13.11. Связь между водным потенциалом листьев и степенью транспирации.
Как правило, линейной связью описывают сумму всех гидравлических сопротивлений в континууме почва — лист (диагональные прямые а, Ь. с обозначают постепенно повышающееся сопротивление транс- локации). Потенциал при транспирации, равной 0, означает равновесное состояние между растением и почвой в ранние утренние часы (PD, predawn water potential). А— нормальное снижение без гистерезиса; В — гистерезис в течение дня (время нанесено на график), указывающий на увеличивающееся затруднение водного транспорта (кавитация ксилемы или обезвоживание почвы вблизи тонких корней). При влажной почве большая крутизна линии регрессии также отражает патологические изменения в ксилеме или повреждения тонких корней (например, поражение грибами или корневая гниль)
лема способна быстро ликвидировать кавитацию.
Отчетливые различия в критических значениях водного потенциала, при которых проявляются первые эмболии (ультразвуковые измерения на засыхающих ветвях) у различных видов, могут коррелировать с диаметром проводящих путей (при большом диаметре эмболия более вероятна), что с другой стороны, возможно, находится в связи со специфической глубиной проникновения корней (до достигаемой почвенной влаги) и ритмами сезонного развития (влажный и сухой периоды). В средиземноморских смешанных насаждениях, состоящих из сезонно-зеленых видов с крупными порами и вечнозеленых с мелкими, последние, которые по сравнению с первыми обладают более глубокими корнями, менее (но более длительно) активны и в сухие периоды менее подвержены эмболии. У видов древесных растений, обитающих в сухих местах, эти признаки ксилемы более выражены.
Еще по теме 13.4. Механические воздействия:
- Почвы, нарушенные механическими воздействиями
- Повреждения механического происхождения.
- ЗАКРЫТЫЕ МЕХАНИЧЕСКИЕ ПОВРЕЖДЕНИЯ
- Механические устройства для гомогенизации
- СУДЕБНО-ВЕТЕРИНАРНАЯ ТРАВМАТОЛОГИЯ. ЭКСПЕРТИЗА ПОВРЕЖДЕНИЙ МЕХАНИЧЕСКОГО ПРОИСХОЖДЕНИЯ
- Другие механические транспортные средства
- МЕХАНИЧЕСКИЕ ФОРМЫ КОЛИК (НЕПРОХОДИМОСТЬ КИШОК - ИЛЕУС)
- Особенности ботанического состава, воднофизических и физико-механических свойств видов торфа
- ВОЗДЕЙСТВИЕ ПАРАЗИТОВ НА ОРГАНИЗМ ХОЗЯИНА
- Виды антропогенных воздействий
- Мониторинг и антропогенные воздействия
- Основные принципы воздействия абиотических факторов
- Воздействие извне
- Воздействие света на насекомых